

Ezjakintasunaren kartografia #157
Izan liteke baraua simulatzen duen dieta bat diabetesaren aurrean erantzuna? Rosa García-Verdugok dakarkigu gaia: Fasting against diabetes.
LIGOk laserrak erabili zituen grabitazio-uhinak detektatzeko, hauen ordez erloju atomikoak erabili daitezke uhinok antzemateko. Victor Marinek kontatzen digu zelan: The gravitational wave detector revisited: a new approach using atomic clocks.
Antibiotikoen aurrean sortutako erresistentzia gero eta arazo larriagoa da. Egoera honen aurrean beharrezkoak dira botika berriak eta, baita ere, egun ditugun antibiotikoak emateko sistema eraginkorragoak. Ildo honetan lanean dihardu DIPC: Combatting antimicrobial resistance with a ruthenium-based photorelease antimicrobial therapy.
–—–
Mapping Ignorance bloga lanean diharduten ikertzaileek eta hainbat arlotako profesionalek lantzen dute. Zientziaren edozein arlotako ikerketen azken emaitzen berri ematen duen gunea da. UPV/EHUko Kultura Zientifikoko Katedraren eta Nazioarteko Bikaintasun Campusaren ekimena da eta bertan parte hartu nahi izanez gero, idatzi iezaguzu.
The post Ezjakintasunaren kartografia #157 appeared first on Zientzia Kaiera.
Ezjakintasunaren kartografia #157

Izan liteke baraua simulatzen duen dieta bat diabetesaren aurrean erantzuna? Rosa García-Verdugok dakarkigu gaia: Fasting against diabetes.
LIGOk laserrak erabili zituen grabitazio-uhinak detektatzeko, hauen ordez erloju atomikoak erabili daitezke uhinok antzemateko. Victor Marinek kontatzen digu zelan: The gravitational wave detector revisited: a new approach using atomic clocks.
Antibiotikoen aurrean sortutako erresistentzia gero eta arazo larriagoa da. Egoera honen aurrean beharrezkoak dira botika berriak eta, baita ere, egun ditugun antibiotikoak emateko sistema eraginkorragoak. Ildo honetan lanean dihardu DIPC: Combatting antimicrobial resistance with a ruthenium-based photorelease antimicrobial therapy.
–—–
Mapping Ignorance bloga lanean diharduten ikertzaileek eta hainbat arlotako profesionalek lantzen dute. Zientziaren edozein arlotako ikerketen azken emaitzen berri ematen duen gunea da. UPV/EHUko Kultura Zientifikoko Katedraren eta Nazioarteko Bikaintasun Campusaren ekimena da eta bertan parte hartu nahi izanez gero, idatzi iezaguzu.
The post Ezjakintasunaren kartografia #157 appeared first on Zientzia Kaiera.
Ciencia, Modernidad y Posmodernidad

En la historia hay edades que más o menos todos ubicamos bien. En lo que se refiere a Europa la Edad Antigua va desde las primeras civilizaciones hasta la caída del Imperio Romano de Occidente en el año 476; la Edad Media desde esta fecha al descubrimiento de América en 1492; la Edad Moderna desde este momento a la Revolución Francesa de 1789; y, desde entonces estaríamos en la Edad Contemporánea. Pero hay otra posibilidad de distinguir eras históricas que se basa en los conceptos de ciencia, tecnología y razón y sus papeles en el proceso histórico: Modernidad y Posmodernidad.

Hegel
El término “moderno” aparece por primera vez en el siglo XVI como una forma de señalar la separación del presente con el pasado. Y aunque en el siglo XVII los filósofos naturales experimentales se distinguían a sí mismos de las prácticas del pasado en el debate entre “antiguos” y “modernos”, fue Georg Wilhelm Friedrich Hegel (1771-1831) quien dio la primera definición formal de modernidad como la persistente re-creación del yo y las condiciones de vida. Desde Hegel existe quien da en llamar Modernidad al período que va de la Revolución Francesa (1789) al final de la Segunda Guerra Mundial (1946). En 1947 los arquitectos comienzan a hablar de la existencia de un estilo “posmoderno” y conforme avanza la Guerra Fría los filósofos y sociólogos articulan el significado (o mejor, significados) de Posmodernidad. Modernos y posmodernos varios coinciden en que la Posmodernidad es la era en la que estamos desde los ochenta.
Tras Hegel la Modernidad se refiere a dos narrativas “maestras”, a saber, el desarrollo de la promesa de la Ilustración de la liberación a través de la razón, y la unificación de todas las ramas del conocimiento mediante la extensión de los métodos racionales de la ciencia y la tecnología. Jürgen Habermas (n. 1929) mantiene que ambas narrativas se han empleado para legitimar la ciencia y la tecnología en el ámbito social desde 1800. La ciencia, para los teóricos de lo moderno, es una práctica representativa, una reproducción de la naturaleza, cuyas características son el resultado del consenso racional entre los que la practican, quienes, a su vez, establecen verdades científicas independientes de los intereses sociales. Desde el momento en que el consenso implica la ausencia de conflicto, desde el punto de vista moderno la ciencia es un instrumento racional de paz.
Hay pensadores, como Habermas o Bruno Latour (n. 1947), que creen que la Modernidad es un proyecto inacabado que aún merece la pena proseguir con objeto de conseguir una sociedad libre de conflictos en la que la comunicación clara y racional es posible. Desde la perspectiva modernista la historia de la ciencia y la tecnología se encuadra claramente dentro de las narrativas teleológicas de progreso, racionalización, secularización, burocratización e, incluso, de estado-nación.

Nietzsche
Los orígenes de la posmodernidad los podríamos trazar hasta la crítica de la razón de Friedrich Nietzsche (1844-1900) y otros a finales del XIX. Pero no será hasta después de las atrocidades del Tercer Reich que se empezó a dudar seriamente del proyecto moderno de liberación y unificación mediante la razón. La Posmodernidad no solo rechaza estas dos narrativas maestras de la Modernidad, sino también la idea de que la representación (tanto en las artes como en las ciencias) pueda ser un espejo preciso de la realidad objetiva.
Los posmodernos como Jean-François Lyotard (1924-1998) reconceptualizan las ciencias como actividades orientadas hacia la reproducción de las prácticas de investigación más que a la producción de resultados acerca del mundo. Lo que Lyotard llama “performatividad” en las ciencias puede apreciarse en la inclinación de las ciencias hacia la interdisciplinaridad y el trabajo en equipo. En la incesante búsqueda de “lo nuevo” (la invención de nuevos vocabularios, prácticas, reglas de investigación, consensos) la ciencia posmoderna es una forma de conocimiento que no reproduce lo conocido, sino que constantemente busca lo desconocido y lo indeterminado.
El carácter anti-representativo de la ciencia posmoderna significa que las imágenes (y las teorías) no representan la naturaleza, sino que son una proyección de los entornos políticos, sociales y económicos en los que se desarrolla la actividad científica. Dicho de otro modo, la naturaleza misma desaparece en un mundo creado de simulacros y simulaciones y la historia de las ciencias se convierte en la historia de las imágenes y de lo que se cree que se encuentra inmerso en ellas.
En la Posmodernidad las ciencias son una parte de la economía, pero en un papel muy diferente al que jugaban en la Modernidad. Ahora sus imágenes son sujetos de la ideología política y, por tanto, su verdad es relativa: frente a unos hechos, existen hechos alternativos. De aquí se sigue la “caridad epistemológica” hacia las creencias irracionales que se encuentra habitualmente entre los posmodernos.
Este post ha sido realizado por César Tomé López (@EDocet) y es una colaboración de Naukas con la Cátedra de Cultura Científica de la UPV/EHU.
El artículo Ciencia, Modernidad y Posmodernidad se ha escrito en Cuaderno de Cultura Científica.
Entradas relacionadas:Josu Lopez-Gazpio: “Homeopatiaren arrakasta plazebo efektuaren bidez azaldu daiteke” #Zientzialari (71)
Gizartean oso hedatua dagoen sendabidea da homeopatia. Hala ere, ugariak dira Kimika, Fisika, Biologia eta Medikuntzatik egiten zaizkion kritikak. Izan ere, zientziaren arloan egin diren ikerketa zorrotz eta kontrolatuek ez dute inoiz frogatu homeopatia plazebo efektua baino eraginkorragoa denik. Zergatik du orduan arrakasta sendabide honek? Zeintzuk dira bere printzipioak? Nola prestatzen dira sendagai homeopatikoak?
Josu Lopez-Gazpio Kimikan doktorea da eta berarekin hitz egin dugu galderen erantzuna bilatzeko. Bere ustez, arduraz jokatu behar da homeopatiarekin, ebidentzian oinarritzen ez den medikuntza arriskutsua izan daitekeelako.
“Zientzialari” izeneko atal honen bitartez zientziaren oinarrizko kontzeptuak azaldu nahi ditugu euskal ikertzaileen laguntzarekin.
The post Josu Lopez-Gazpio: “Homeopatiaren arrakasta plazebo efektuaren bidez azaldu daiteke” #Zientzialari (71) appeared first on Zientzia Kaiera.
Josu Lopez-Gazpio: “Homeopatiaren arrakasta plazebo efektuaren bidez azaldu daiteke” #Zientzialari (71)
Gizartean oso hedatua dagoen sendabidea da homeopatia. Hala ere, ugariak dira Kimika, Fisika, Biologia eta Medikuntzatik egiten zaizkion kritikak. Izan ere, zientziaren arloan egin diren ikerketa zorrotz eta kontrolatuek ez dute inoiz frogatu homeopatia plazebo efektua baino eraginkorragoa denik. Zergatik du orduan arrakasta sendabide honek? Zeintzuk dira bere printzipioak? Nola prestatzen dira sendagai homeopatikoak?
Josu Lopez-Gazpio Kimikan doktorea da eta berarekin hitz egin dugu galderen erantzuna bilatzeko. Bere ustez, arduraz jokatu behar da homeopatiarekin, ebidentzian oinarritzen ez den medikuntza arriskutsua izan daitekeelako.
“Zientzialari” izeneko atal honen bitartez zientziaren oinarrizko kontzeptuak azaldu nahi ditugu euskal ikertzaileen laguntzarekin.
The post Josu Lopez-Gazpio: “Homeopatiaren arrakasta plazebo efektuaren bidez azaldu daiteke” #Zientzialari (71) appeared first on Zientzia Kaiera.
Cuando los científicos marchan

El ejercicio de la ciencia es una metódica aproximación al uso más puro posible de la razón. En cada paso del método científico se intenta, con deliberación y cuidado, eliminar el más mínimo atisbo de emoción, de apego a las ideas propias, de rechazo de las ajenas basado en antipatías personales; se dan los pasos pertinentes para que sean los datos y no las emociones las que decidan. Y se fracasa, claro, porque los científicos son humanos; porque el cerebro humano en su propio modo de funcionamiento tiene la razón enredada con la emoción, los sentimientos creciendo como hiedras sobre los datos (o viceversa). En eso, ay, quienes se dedican a la ciencia no se diferencias del resto de sus compañeros de especie, y no pueden diferenciarse. Los científicos no son internamente ‘más racionales’ que los demás.
Pero lo intentan. Precisamente por eso existe el método científico: porque si quieres conocer la naturaleza con la mayor precisión posible es necesario filtrar, todo lo que se pueda, los factores emocionales. Para eso se hacen test de doble ciego, complejos diseños experimentales o enrevesados cálculos estadísticos. Por eso los artículos se codifican en un lenguaje artificial y preciso y se revisan por pares anónimos antes de salir publicados. Por eso los tribunales de tesis, y las solicitudes de proyectos con evaluadores externos. La diferencia entre los practicantes de ciencia y el resto de los mortales está en que los científicos son conscientes de las limitaciones de su propio conocimiento y luchan enconada y constantemente por eliminar las fuentes de conflicto entre emoción y razón. Es una lucha condenada a la derrota, porque ningún científico humano puede dejar de ser humano. Pero gracias a ella cada vez conocemos más hechos objetivos sobre el cosmos, lo cual es bello en sí mismo y además ha resultado ser enormemente útil a nuestra especie.
Por eso la ciencia es necesaria y automáticamente enemiga de cualquier doctrina religiosa o política que se base sobre todo en la emoción. Por la sencilla razón de que su método y objetivo final son incompatibles con las llamadas a guiarse por los sentimientos y abandonar la razón. Con independencia de las ideologías políticas o religiosas de cada cual la ciencia siempre se transforma en una muralla dirigida contra ciertas formas de religión o política: aquellas que prefieren fomentar sentimientos por encima de las realidades, las que agitan emociones sin tener en cuenta los datos. No porque la propia actividad científica tenga un sesgo político concreto, sino porque por definición la ciencia y quienes la practican están del lado de la razón y los hechos, y por tanto en contra de quienes se enfrentan al raciocinio o ignoran los datos para respaldar sus ideologías.
Éste es el verdadero trasfondo de las Marchas por la Ciencia del pasado 22 de abril de 2017, y la razón por la que se ha acusado a sus participantes de convertir a la ciencia en una pieza más del tablero político. Cosa que podrían haber hecho, legítimamente: determinadas opciones políticas han agredido a la actividad científica profesional a través de recortes de presupuestos, reducciones de plantillas, instituciones paralizadas y otras acciones que en general han resultado en menos sueldos, menos proyectos, mayores dificultades y menos futuro profesional. Los científicos, como los mineros, los médicos o los conductores de camión, tienen derecho a protestar cuando sus condiciones profesionales se deterioran como cualquier otro tipo de trabajador.
Aunque el impulso que ha sacado a los científicos a las calles de centenares de ciudades en todo el mundo va más allá; mucho más allá: se trata de una defensa del conocimiento y del raciocinio de la humanidad frente a fuerzas que pretenden llevarnos de vuelta a una concepción del cosmos en la que lo único que importa son las pasiones humanas y no la realidad. Se trata de defender la toma de decisiones que tiene en cuenta los datos; la existencia de una realidad empírica que no se puede obviar por mucho que se desee políticamente; el papel del conocimiento y la razón en los procesos que definen el futuro de las naciones. Quienes rechazan los datos y acusan de sesgada y comprometida a la ciencia pretenden que nadie oponga realidades a sus manipulaciones emocionales; que ni siquiera la realidad imponga límites a su voluntad, expresada en términos de pasión de las masas. Conocemos a dónde lleva este camino; ya lo hemos recorrido antes, siempre con terroríficos resultados.
La cultura gremial de quienes practican ciencia e incluso sus querencias personales no son las más dadas a la protesta pública que existen. De hecho por temperamento y por práctica diaria los científicos con meticulosos hasta la pedantería, extremadamente precisos en lenguaje y afirmaciones, críticos de cualquier detalle propio y ajeno, solitarios y poco dados a formar turbas. La comunidad científica tiene una cuota mayor de lo común de raros, obsesos, detallistas y tocanarices; los científicos muy raras veces están de acuerdo en algo, y de esas aún menos tanto como para salir a la calle en grandes prupos y exteriorizar su protesta. Esta vez ha sido así porque muchos de ellos sienten que la apuesta es mayor de lo normal; que el peligro va más allá de la financiación y los detalles de las becas o proyectos y afecta al futuro mismo de la sociedad. Una pancarta en una de las manifestaciones estadounidenses decía “So bad even the introverts are here’ (tan mal estamos que hasta los introvertidos estamos aquí). Porque cuando los científicos marchan es que el peligro es grande, y real.
Sobre el autor: José Cervera (@Retiario) es periodista especializado en ciencia y tecnología y da clases de periodismo digital.
El artículo Cuando los científicos marchan se ha escrito en Cuaderno de Cultura Científica.
Entradas relacionadas:- NASA, ¿cuándo vas a llevarnos a Venus en un barco (o en lo que sea)?
- ¿Son arrogantes los científicos?
- Artistas científicos
Galdutako espezieen bila
Gizakiaren esku-hartzeak sekulako kaltea egin dio gure planetaren biodibertsitateari. Habitat ugari suntsitu ditugu zuzenean zein zeharka, eta horietan bizi ziren eta egoera berrira egokitzen asmatu ez duten makina bat espezie desagertu da. Hor ez dago atzera bueltarik. Badira, ordea, desagertuta ez, baina galduta dauden espezieak: adituek ez dituzte ikusi aspaldian (batzuetan, batere irisgarriak ez diren tokietan bizi direlako), baina ale gutxi izan arren, badaude zantzuak oraindik badirela pentsatzeko.
Haientzat ere beranduegi izan baino lehen, galdutako espezieok berraurkitzeko kanpaina abiarazi du Global Wildlife Conservation (GWC) erakundeak. The Search for Lost Species (galdutako espezieen bila) du izena ekimenak. Ehun zientzialari baino gehiagorekin elkarlanean, 160 estatutan galdutzat jotzen diren 1.200 animalia eta landare espezieren zerrenda egin dute: gutxienez azken hamarkadan zientzialari bakar batek ere ez ditu ikusi. Gainera, horien artetik, 25 desiratuenak aukeratu dituzte, eta azken horiexek aurkitzea da lehentasuna.
{kind=link}

1. irudia: Jomugan jarri dituzten 25 espezie nagusiak irudikatzen dituen kartela)
(Argazkia: Alexis Rockman / Global Wildlife Conservation)
Robin Moore GWCko komunikazio zuzendariak adierazi bezala, “animalia eta landare xelebre eta karismatikoak daude espezie horien artean, eta kontserbaziorako egundoko aukera dakarte. Espezie iheskor horietako edozein berraurkitzeak haren misterioak askatzen lagunduko liguke, eta informazio baliagarria emango liguke espeziea, bere habitata eta inguruko fauna ulertu eta hobeto kontserbatzeko. Ahaztutako espezie horietako askoren desagertzea galarazteko azken aukera izan liteke hau”.
Lehentasuntzat jo dituzten 25 espezie desiratuenek 18 estatutan dute jatorria, eta horien artean daude hamar ugaztun; hiruna hegazti, narrasti eta arrain; bi anfibio; eta intsektu, krustazeo, koral eta landare bana. Espezie horietako zazpiren kasuan, gutxienez, azken hogei urteotan ikusiak izan direla dirudi, baina oraingoz zientzialariek ezin izan dute haien presentzia berretsi. Bestalde, 25 espezie horietako batzuk aurkitzea erronka bereziki zaila izango dela dirudi. Izan ere, zenbaiten kasuan, ale bakarra baino ez dute antzeman ikertzen hasi zirenetik, eta duela hainbat hamarkada, gainera.

2. irudia: Buru arrosadun ahatearen ale disekatu bat, Eskoziako Museo Nazionalean. 1949an ikusi zuten azken aldiz, Indian) (Argazkia: Geni / GFDL CC-BY-SA)
Aniztasuna izan dute helburu 25 espezie horiek aukeratzean: ingurune eta mota oso desberdinetakoak dira. Esaterako, Wondiwoi zuhaitz kangurua 1928an ikusi zuten azken aldiz, Indonesian. Buruan luma arrosa distiratsuak dituen ahate espezie bat 1949an erregistratu zuten azkenekoz, Indian. Zerrendan badago Galapagoetako Fernandina uhartean baino aurkitu ez duten dortoka espezie bat ere, eta 1906tik ez dute ikusi, gainera; baina egiari zor, oso irla basatia da eta ez da batere irisgarria, eta orain arte ez dute nahi bezainbeste ikertu, beraz. Antzeko zerbait gertatzen da Australiako itsas zaldi espezie oso txiki batekin ere: bizitzekotan, gizakiarentzat batere eskuragarri ez dagoen ingurune batean bizi da.
Espezie horietako edozein berraurkitzeak garrantzi handia izango luke biodibertsitatearen ikuspegi soilari begiratuta, baina ez horregatik bakarrik. Venezuelako apo arlekin eskarlata topatzea, adibidez, lagungarria izango litzateke kitridoekiko erresistentzia hobeto ulertzeko, onddo patogeno horiek anfibioak akabatzen ari baitira, munduan luze-zabal.
Kanpainaren lehen fase honetan dirua biltzen dihardu GWC erakundeak. Guztira milioi erdi dolar eskuratu nahi dute, galdutako espezie horiek bizi omen diren inguruneetan espedizio zehatzak egin, eta banan-banan, horietako bakoitza berraurkitzen ahalegintzeko.
Kanpainaren berri ematen duen bideoa, ingelesez
———————————————————————————-
Egileaz: Amaia Portugal (@amaiaportugal) zientzia kazetaria da.
———————————————————————————
The post Galdutako espezieen bila appeared first on Zientzia Kaiera.
Galdutako espezieen bila
Gizakiaren esku-hartzeak sekulako kaltea egin dio gure planetaren biodibertsitateari. Habitat ugari suntsitu ditugu zuzenean zein zeharka, eta horietan bizi ziren eta egoera berrira egokitzen asmatu ez duten makina bat espezie desagertu da. Hor ez dago atzera bueltarik. Badira, ordea, desagertuta ez, baina galduta dauden espezieak: adituek ez dituzte ikusi aspaldian (batzuetan, batere irisgarriak ez diren tokietan bizi direlako), baina ale gutxi izan arren, badaude zantzuak oraindik badirela pentsatzeko.
Haientzat ere beranduegi izan baino lehen, galdutako espezieok berraurkitzeko kanpaina abiarazi du Global Wildlife Conservation (GWC) erakundeak. The Search for Lost Species (galdutako espezieen bila) du izena ekimenak. Ehun zientzialari baino gehiagorekin elkarlanean, 160 estatutan galdutzat jotzen diren 1.200 animalia eta landare espezieren zerrenda egin dute: gutxienez azken hamarkadan zientzialari bakar batek ere ez ditu ikusi. Gainera, horien artetik, 25 desiratuenak aukeratu dituzte, eta azken horiexek aurkitzea da lehentasuna.
 1. irudia: Jomugan jarri dituzten 25 espezie nagusiak irudikatzen dituen kartela)
1. irudia: Jomugan jarri dituzten 25 espezie nagusiak irudikatzen dituen kartela)(Argazkia: Alexis Rockman / Global Wildlife Conservation)
Robin Moore GWCko komunikazio zuzendariak adierazi bezala, “animalia eta landare xelebre eta karismatikoak daude espezie horien artean, eta kontserbaziorako egundoko aukera dakarte. Espezie iheskor horietako edozein berraurkitzeak haren misterioak askatzen lagunduko liguke, eta informazio baliagarria emango liguke espeziea, bere habitata eta inguruko fauna ulertu eta hobeto kontserbatzeko. Ahaztutako espezie horietako askoren desagertzea galarazteko azken aukera izan liteke hau”.
Lehentasuntzat jo dituzten 25 espezie desiratuenek 18 estatutan dute jatorria, eta horien artean daude hamar ugaztun; hiruna hegazti, narrasti eta arrain; bi anfibio; eta intsektu, krustazeo, koral eta landare bana. Espezie horietako zazpiren kasuan, gutxienez, azken hogei urteotan ikusiak izan direla dirudi, baina oraingoz zientzialariek ezin izan dute haien presentzia berretsi. Bestalde, 25 espezie horietako batzuk aurkitzea erronka bereziki zaila izango dela dirudi. Izan ere, zenbaiten kasuan, ale bakarra baino ez dute antzeman ikertzen hasi zirenetik, eta duela hainbat hamarkada, gainera.
 2. irudia: Buru arrosadun ahatearen ale disekatu bat, Eskoziako Museo Nazionalean. 1949an ikusi zuten azken aldiz, Indian) (Argazkia: Geni / GFDL CC-BY-SA)
2. irudia: Buru arrosadun ahatearen ale disekatu bat, Eskoziako Museo Nazionalean. 1949an ikusi zuten azken aldiz, Indian) (Argazkia: Geni / GFDL CC-BY-SA) Aniztasuna izan dute helburu 25 espezie horiek aukeratzean: ingurune eta mota oso desberdinetakoak dira. Esaterako, Wondiwoi zuhaitz kangurua 1928an ikusi zuten azken aldiz, Indonesian. Buruan luma arrosa distiratsuak dituen ahate espezie bat 1949an erregistratu zuten azkenekoz, Indian. Zerrendan badago Galapagoetako Fernandina uhartean baino aurkitu ez duten dortoka espezie bat ere, eta 1906tik ez dute ikusi, gainera; baina egiari zor, oso irla basatia da eta ez da batere irisgarria, eta orain arte ez dute nahi bezainbeste ikertu, beraz. Antzeko zerbait gertatzen da Australiako itsas zaldi espezie oso txiki batekin ere: bizitzekotan, gizakiarentzat batere eskuragarri ez dagoen ingurune batean bizi da.
Espezie horietako edozein berraurkitzeak garrantzi handia izango luke biodibertsitatearen ikuspegi soilari begiratuta, baina ez horregatik bakarrik. Venezuelako apo arlekin eskarlata topatzea, adibidez, lagungarria izango litzateke kitridoekiko erresistentzia hobeto ulertzeko, onddo patogeno horiek anfibioak akabatzen ari baitira, munduan luze-zabal.
Kanpainaren lehen fase honetan dirua biltzen dihardu GWC erakundeak. Guztira milioi erdi dolar eskuratu nahi dute, galdutako espezie horiek bizi omen diren inguruneetan espedizio zehatzak egin, eta banan-banan, horietako bakoitza berraurkitzen ahalegintzeko.
Kanpainaren berri ematen duen bideoa, ingelesez
———————————————————————————-
Egileaz: Amaia Portugal (@amaiaportugal) zientzia kazetaria da.
———————————————————————————
The post Galdutako espezieen bila appeared first on Zientzia Kaiera.
Un algoritmo genético para buscar materiales 2D
Las posibilidades que da la computación para la ciencia, sobre todo los ordenadores potentes pero para nada extraordinarios, es que permiten hacer predicciones que hace solo unos años serían impensables ni siquiera planteárselas. Una reciente investigación en busca de nuevos materiales 2D de uso en nanoelectrónica de un equipo encabezado por Arunima Singh del Instituto Nacional de Estándares y Tecnología de los Estados Unidos es un ejemplo estupendo de este uso.
El grafeno es una forma de carbono muy conocida. Sabemos que es una monocapa plana de átomos de carbono con propiedades muy interesantes. Bien, una de las cosas que demuestra el grafeno es que un material por lo demás ordinario, recordemos que la mina de un lápiz está hecha de láminas de grafeno unas encima de otras formando lo que llamamos grafito, se vuelve extraordinario si lo pasamos de 3 dimensiones a 2 dimensiones.
Efectivamente, algunas propiedades como la magnetorresistencia o la piezoelectricidad solo aparecen en las versiones 2D de los materiales. Entonces, ¿por qué no realizar una búsqueda sistemática de compuestos 2D de los materiales que son del grupo del carbono en la tabla periódica? Recordemos que los elementos de un mismo grupo tienen un comportamiento químico similar porque su estructura electrónica más externa es idéntica.
Antiguamente habría que haber hecho miles de cálculos a mano para intentar averiguar con poca precisión que compuestos 2D serían factible y cuáles podría tener propiedades diferentes. Hoy, lo que estos investigadores han hecho es crear un algoritmo que funciona eliminando todas las variaciones que no sean óptimas dese el punto de vista de estabilidad en un análisis de todas las estructuras 2D hipotéticas de todos los óxidos de cuatro elementos del grupo del carbono: silicio (Si), germanio (Ge), estaño (Sn) y plomo (Pb). Una vez que las tienen calculan las características electrónicas.

Pero esto es aún más complicado de lo que parece. A diferencia del carbono, cuya estructura cristalina 2D (grafeno) es la misma que la 3D (grafito), lo normal es que las estructuras 2D sean diferentes a las 3D. Así, por ejemplo, el cuarzo alfa (estructura 3D del dióxido de silicio, SiO2) tiene una estructura trigonal, pero la 2D, sintetizada recientemente, se ha comprobado que es bitetraédrica.
El algoritmo desarrollado por los investigadores es de los llamados genéticos, una clasificación del campo de la inteligencia artificial que engloba a los inspirados en la evolución biológica y su base genético-molecular. Lo que hace es tomar la población inicial de todos los óxidos posibles y dejarla evolucionar hasta que se encuentran las estructuras de menor nivel de energía y, por tanto, más estables; de éstas solo algunas serán 2D.
Los resultados indican que la forma estable del dióxido de germanio GeO2 debería ser monoclínica, mientras que los dióxidos de estaño SnO2 y plomo PbO2 2D deberían ser 1T (triclínicos, 1 capa), a diferencia de sus versiones 3D que tienen una estructura común de rutilo.
Cálculos adicionales muestran que estas estructuras deberían de poder sintetizarse en el laboratorio, bien sobre sustratos o, como el grafeno, como capas independientes. Los óxidos de germanio y estaño 2D podrían tener aplicaciones interesantes como dieléctricos en transistores mientras que el de estaño podría servir de protección en dispositivos nanoelectrónicos. Ya solo queda sintetizarlos y comprobarlo.
Referencia:
Arunima Singh et al (2017) Genetic algorithm prediction of two-dimensional group-IV dioxides for dielectrics Physical Review B doi: 10.1103/PhysRevB.95.155426
Sobre el autor: César Tomé López es divulgador científico y editor de Mapping Ignorance
Este texto es una colaboración del Cuaderno de Cultura Científica con Next
El artículo Un algoritmo genético para buscar materiales 2D se ha escrito en Cuaderno de Cultura Científica.
Entradas relacionadas:- Siligrafeno, ¿el material para la revolución optoelectrónica?
- Usando ADN para sintetizar nanoestructuras de oro
- Un algoritmo para una resucitación eficiente
Antibiotikoak, bakterioak eta superbakterioak
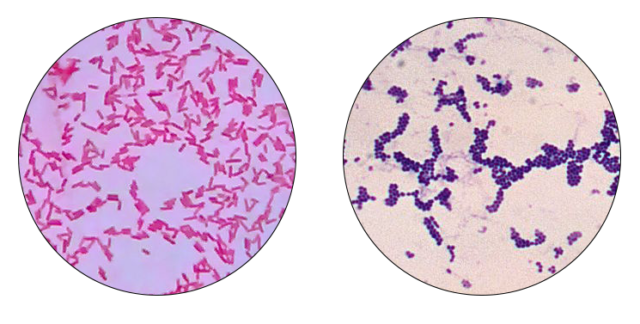
1. irudia: Gram negatibo (ezk.) eta gram positiboak (esk.). Microrao-ren irudian oinarritua (gram negatiboa).
Bakterioek sortutako substantziak dira antibiotikoak, beste bakterioak eraso eta hiltzeko. Bakterioak isolaturik bizi ez direlako, komunitateetan baizik, eta baliabideak eskuratzeko lehiatzen direlako gertatzen da fenomenoa. Modu berean, zenbait bakteriok substantzia horietatik denfendatzeko mekanismoak garatu dituzte.
Antibiotikoei aurre egiteko gaitasuna erresistentzia du izena eta mekanismo hau garatu duten bakterioak ‘superbakterio’. Osasun arloan lehen mailako arazo bihurtu da bakterioen erresistentzia, bereziki bakterio patogenoena (gaixotasunak sortzen dituztenak), Munduko Osasun Erakundeak ere lehentasun gisa ezarri du bakterioen erresistentziaren aurkako borroka.
Ezinbesteko mikroorganismoakGaixotasunak sor ditzaketen mikroorganismoak baino gehiago dira bakterioak. Toki guztietan aurki daitezke eta bizitzarako ezinbestekoak dira, elikagaietan (esnekietan, adibidez) edota hesteetan aurki daitezkeenak, adibidez.
Lurrean 3.500 milioi urte daramaten zelula prokariotoak dira bakterioak. Prokariota izatea nukleo zelularrik ez izatea esan nahi du; kromosomak zitosolean aske izatea, mintzez inguraturik egon gabe. Geneak, beraz, DNA molekula batean daude zitosolean.
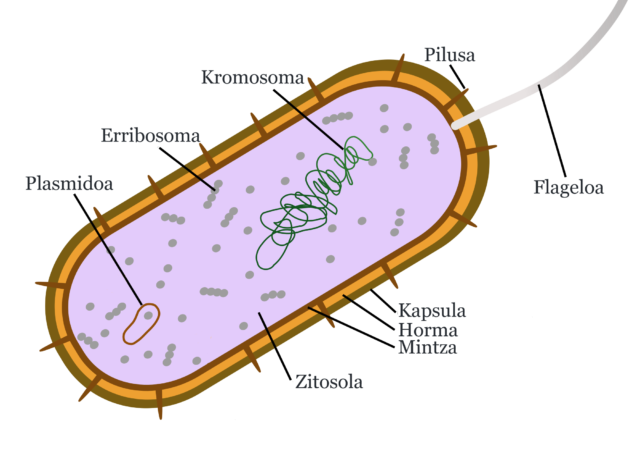
2. irudia: Bakterioaren eskema sinplifikatua. JrPol-en irudian oinarritua. (Egilea: Ziortza Guezuraga)
Oso organulu gutxi izatea da bakterioen beste ezaugarrietako bat, organulu garrantzitsuenak hurrengoak direla:
- Erribosomak. Proteinen sintesia dute funtzio nagusia, kromosoman dauden geneak proteina bilakatzea.
- Mintza. Bi bakterio mota ezberdintzen dira mintzaren arabera, Gram tindaketa baliatuta: Gram + (bi estalkidun mintza dutenak) eta Gram – (hiru estalkidun mintza dutenak).
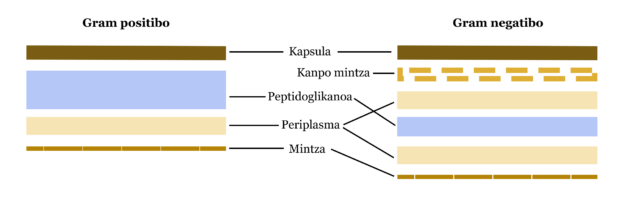
3. irudia: Gram positiboen eta gram negatiboen konparaketa. (Egilea: Ziortza Guezuraga)
- Plasmidoak. DNA molekula txiki zirkularrak dira. Ez dira ezinbestekoak bakterioarentzat, baina dituzten zenbait genek ezaugarri bereziak dituzte ingurune kaltegarrietan bizitzeko, antibiotikoei aurre egiteko, adibidez. Plasmidoetan dute, beraz, erresistentzia bakterioak.
Bakterioak erasotzen dituzten substantziak dira antibiotikoak, bakterioak hil edota haz daitezen eragozten dituztenak. Gizakien kasuan, bakterio patogenoei, gaixotasunak sortzen dituzten bakterioei, aurre egiteko tresna dira. Hiru arlotan erasotzen dute antibiotikoek:
- Estalkiari eragiten. Bakterioen horma zelularrari edota mintzari eragiten diote.
- Proteinen sintesian. Proteinen sintesia eragotzi edota proteina akasdunak sorraraztea bultzatzen dute.
- DNA eta RNA erreplikazioan. DNA eta RNAren erreplikazioa ekiditen dute.
Esan bezala, bakterioak komunitatean bizi dira eta baliabideak lortzeko lehian daude. Logikoa dirudi biziraupenerako lehia horretan zenbait bakteriok gailentzeko eraso teknikak baliatzea. Logikoa da, baita, erantzun moduan defentsak garatzea.
Hiru mekanismo nagusi darabilte bakterioak erresistente bihurtzeko:
- Antibiotikoak degradatu. Antibiotikoa eraldatu egiten dute entzimak erabilita eraginkorra izan ez dadin.
- Efflux. Substantziak zelulatik kanporatzen dituzten ponpak sortzen dituzte.
- Ituen aldaketa. Ituen ezaugarriak aldatzen dituzte antibiotikoak erasotu ez ditzan.
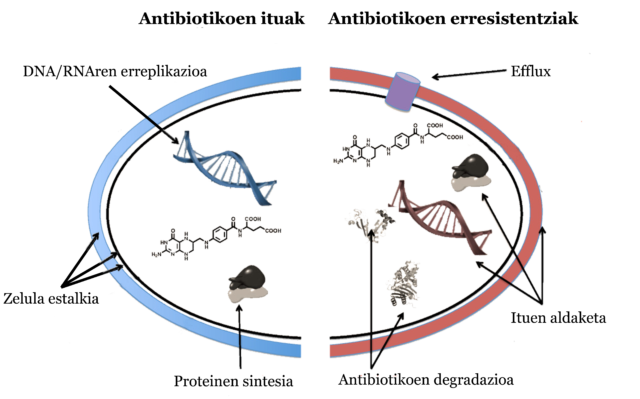
4. irudia: Antibiotikoen ituak eta bakterioen erresistentzia nagusiak. Gerard D Wright-en irudian oinarritua. (Egilea: Ziortza Guezuraga)
Erresistentzia eskuratzenDefentsa mekanismo horiek dira, hain zuzen, erresistentzia. Modu naturalean ematen den fenomenoa da, beraz. Naturala izanik ere, oso kontuan izateko bi faktore ditu erresistentziak: Antibiotikoen erabilera okerra eta erresistentzien zabalkuntza. Azken hau da bereziki kezkagarria, antibiotikoei aurre egiteko estrategiak bakterioz bakterio hedatzen dira, erresistentzia zabaltzen dela. Bi modu nagusitan gertatzen da erresistentziaren zabalkuntza:
Populazioaren biziraupena. Antibiotikoek hiltzen ez dituzten populazioak hazi egiten dira, bakterio erresistenteen biziraupena bermatzen duena.
Geneen transferentzia horizontala. Erresistente bilakatzen dituzten geneak bakterioz bakterio pasatzea. Hiru transferentzia mota daude:
1. Transformazioa. A bakterio batek DNA askatzen du mediora eta B Bakterioak DNA molekulak hartzen ditu mediotik, zuzenean hormatik zehar sartzen direnak.
5. irudia: Transformazioaren prozesua. (Egilea: Ziortza Guezuraga)
2. Transdukzioa. Fagoak (birusak) baliatuta bakteria batetik bestera eramaten da DNA.
6. irudia: Transdukzioaren prozesua. (Egilea: Ziortza Guezuraga)
3. Konjugazioa. Bakterioen arteko kontaktu zuzenaren bidez egiten da transferentzia. Lehen aipatutako plasmidoak baliatuta geneak bakterio batetik bestera pasatzen dira.
Konjugazioa gertatzeko plasmido konjugatiboak behar dira, erresistentzia sortzen duten geneez gain, transferentzia aktibatzeko geneak ere dituztenak. Hiru pausutan ematen da:
- Bakterio emaileak pilusa sortzen du transferentzia geneei esker. Pilusa bi bakterioen arteko lotura izango da, pasabidea.
- Bi bakterioak elkartzean plasmidoa aldatu egiten da eta erlaxosoma bihurtzen da. Kate bikoitza dute plasmidoek eta horretako harizpi bat bakterio emailetik hartzailera pasatzen da.
- Bai bakterio emaileak bai hartzaileak harizpia kate bikoitz bihurtzen dute. Modu honetan, gene erresistenteak eta transferentzia geneak eskuratuta, emaile bilakatzen da ordura arte hartzailea izan den bakterioa.
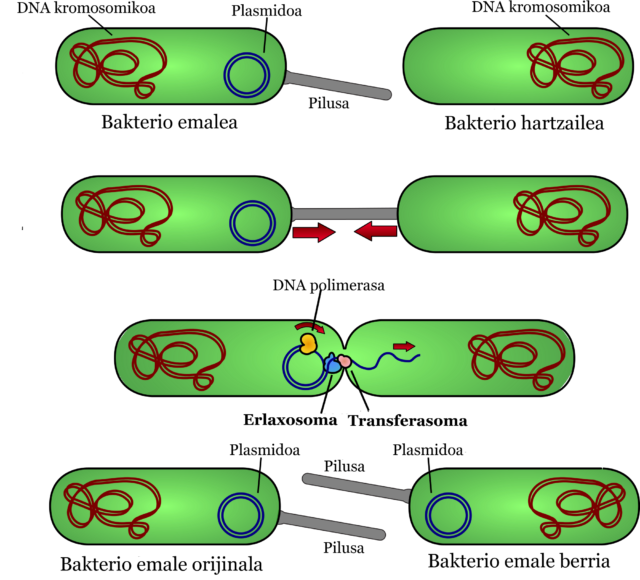
7. irudia: Konjugazioaren prozesua. (Egilea: Ziortza Guezuraga)
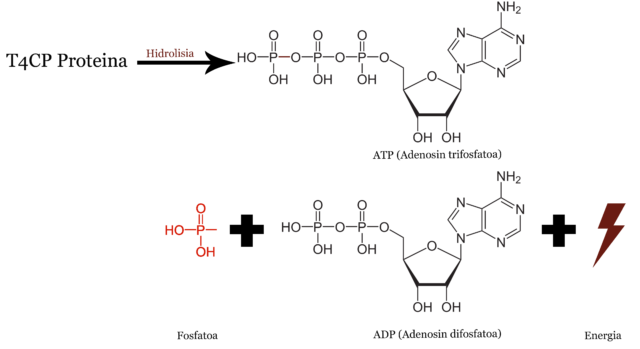
Proteina motorraren bilaKonjugazioa da, hain justu, Itziar Alkorta eta bere taldea ikertzen ari diren mekanismoa. Labur esanda, plasmidoa bakterio batetik bestera pasatzea ahalbidetzen duen proteina, T4CP izenekoa, ezeztatzea dute helburu. Akopladorea eta motor molekularra da T4CP proteina: ATP molekula hidrolizatzen du ADPa eta energia askatuz, eta askatzen den energia hori DNA harizpia beste bakteriora ponpatzeko erabiltzen du zelulak.
8.irudia: “Superbakterioek” sortzen dituzten infekzioak osasun publikoko arazo larria bihurtzen ari dira, Bakterioen konjugazioa da antibiotikoen aurrean bakterioek azaleratzen duten barreiaduraren erantzulea. Prozesu horretan, batez ere, T4CP proteina aklopadorea da funtsezkoa. (Egilea: Ziortza Guezuraga)
Konjugazioaren erdian dago proteina eta funtsezkoa da prozesua gerta dadin, T4CP proteinaren faltan bakterio konjugazioa ez da gertatzen. Mintz proteina da T4CP, ikertzeko zailak direnak, challenging proteins bezala ezagunak dira. Lan egiteko oso zailak dira, oso baldintza zehatzak behar dituzte eta datuak lortzea asko kostatzen da. 20 urte daramatzate proteina hau ikertzen.
Nahiz eta hasiera batean T4CP proteina baino ez zuten aztertzen, ikerketa taldeak hainbat proteina akopladore aztertzen ditu egun, ezaugarri komunak identifikatzeko. Batez ere, ATParen erabilerari dagokionez.
Teknika biokimiko, biofisiko eta genetikoak erabiltzen dituzte proteina hauek zelan funtzionatzen diren ikertzeko. Patroiak aurkitu eta inhibizioa lortzeko estrategia komuna garatu daiteke proteina akopladore guztietarako. Konjugazioa kontrolatu ahal izateko. Horretaz gain, alderdi aplikatua ere badu ikerketak.
- Kimiotekak aztertzen ari dira, bertako konposatuak aztertuta T4PC proteina inhibitzeko baliagarriak izan daitezkeenak aurkitzeko, hauen bidez bakterio batetik besterako transferentzia kontrolatzeko.
- Bilduma teorikoak ere erabiltzen ari dira. Kimiotekiko ezberdintasuna da inhibitzaile posibleak teorikoki aztertzen direla, proteinaren ezaugarriak kontuan izanda.
Bakterioen aurkako borrokan antibiotikoek egiten duten eraso zuzenarekin batera erresistentzien hedapenaren kontrola konbinatzea da ikerketaren helburua. Infekzioak kontrolatzeko antibiotikoak erabili behar dira, baina konjugazio inhibitzaileekin konbinatuta, erresistentzia eta erresistentziaren hedapena ekidin daitezke.
Bakterio infekziosoen aurkako borrokan, beraz, garrantzitsua da antibiotikoak topatu eta garatzeaz gain, bestelako estrategiekin konbinatzea. Sagua eta katua bezala, antibiotiko berriak sortzen zientziak eta erresistentziak garatzen bakterioak, ibili beharrean irtenbide definitiboagoak lortzeko.
Artikulu hau Itziar Alkortak Azkuna Zentroan emandako hitzaldian dago oinarritua:
———————————————————————–
Egileaz: Ziortza Guezuraga kazetaria eta UPV/EHUko Kultura Zientifikoko Katedraren kolaboratzailea da.
———————————————————————–
The post Antibiotikoak, bakterioak eta superbakterioak appeared first on Zientzia Kaiera.
Antibiotikoak, bakterioak eta superbakterioak
 1. irudia: Gram negatibo (ezk.) eta gram positiboak (esk.). Microrao-ren irudian oinarritua (gram negatiboa).
1. irudia: Gram negatibo (ezk.) eta gram positiboak (esk.). Microrao-ren irudian oinarritua (gram negatiboa). Bakterioek sortutako substantziak dira antibiotikoak, beste bakterioak eraso eta hiltzeko. Bakterioak isolaturik bizi ez direlako, komunitateetan baizik, eta baliabideak eskuratzeko lehiatzen direlako gertatzen da fenomenoa. Modu berean, zenbait bakteriok substantzia horietatik denfendatzeko mekanismoak garatu dituzte.
Antibiotikoei aurre egiteko gaitasuna erresistentzia du izena eta mekanismo hau garatu duten bakterioak ‘superbakterio’. Osasun arloan lehen mailako arazo bihurtu da bakterioen erresistentzia, bereziki bakterio patogenoena (gaixotasunak sortzen dituztenak), Munduko Osasun Erakundeak ere lehentasun gisa ezarri du bakterioen erresistentziaren aurkako borroka.
Ezinbesteko mikroorganismoakGaixotasunak sor ditzaketen mikroorganismoak baino gehiago dira bakterioak. Toki guztietan aurki daitezke eta bizitzarako ezinbestekoak dira, elikagaietan (esnekietan, adibidez) edota hesteetan aurki daitezkeenak, adibidez.
Lurrean 3.500 milioi urte daramaten zelula prokariotoak dira bakterioak. Prokariota izatea nukleo zelularrik ez izatea esan nahi du; kromosomak zitosolean aske izatea, mintzez inguraturik egon gabe. Geneak, beraz, DNA molekula batean daude zitosolean.
 2. irudia: Bakterioaren eskema sinplifikatua. JrPol-en irudian oinarritua. (Egilea: Ziortza Guezuraga)
2. irudia: Bakterioaren eskema sinplifikatua. JrPol-en irudian oinarritua. (Egilea: Ziortza Guezuraga) Oso organulu gutxi izatea da bakterioen beste ezaugarrietako bat, organulu garrantzitsuenak hurrengoak direla:
- Erribosomak. Proteinen sintesia dute funtzio nagusia, kromosoman dauden geneak proteina bilakatzea.
- Mintza. Bi bakterio mota ezberdintzen dira mintzaren arabera, Gram tindaketa baliatuta: Gram + (bi estalkidun mintza dutenak) eta Gram – (hiru estalkidun mintza dutenak).
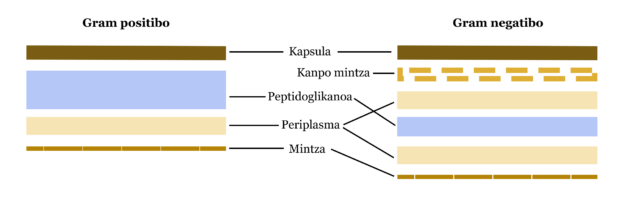 3. irudia: Gram positiboen eta gram negatiboen konparaketa. (Egilea: Ziortza Guezuraga)
3. irudia: Gram positiboen eta gram negatiboen konparaketa. (Egilea: Ziortza Guezuraga) - Plasmidoak. DNA molekula txiki zirkularrak dira. Ez dira ezinbestekoak bakterioarentzat, baina dituzten zenbait genek ezaugarri bereziak dituzte ingurune kaltegarrietan bizitzeko, antibiotikoei aurre egiteko, adibidez. Plasmidoetan dute, beraz, erresistentzia bakterioak.
Bakterioak erasotzen dituzten substantziak dira antibiotikoak, bakterioak hil edota haz daitezen eragozten dituztenak. Gizakien kasuan, bakterio patogenoei, gaixotasunak sortzen dituzten bakterioei, aurre egiteko tresna dira. Hiru arlotan erasotzen dute antibiotikoek:
- Estalkiari eragiten. Bakterioen horma zelularrari edota mintzari eragiten diote.
- Proteinen sintesian. Proteinen sintesia eragotzi edota proteina akasdunak sorraraztea bultzatzen dute.
- DNA eta RNA erreplikazioan. DNA eta RNAren erreplikazioa ekiditen dute.
Esan bezala, bakterioak komunitatean bizi dira eta baliabideak lortzeko lehian daude. Logikoa dirudi biziraupenerako lehia horretan zenbait bakteriok gailentzeko eraso teknikak baliatzea. Logikoa da, baita, erantzun moduan defentsak garatzea.
Hiru mekanismo nagusi darabilte bakterioak erresistente bihurtzeko:
- Antibiotikoak degradatu. Antibiotikoa eraldatu egiten dute entzimak erabilita eraginkorra izan ez dadin.
- Efflux. Substantziak zelulatik kanporatzen dituzten ponpak sortzen dituzte.
- Ituen aldaketa. Ituen ezaugarriak aldatzen dituzte antibiotikoak erasotu ez ditzan.
 4. irudia: Antibiotikoen ituak eta bakterioen erresistentzia nagusiak. Gerard D Wright-en irudian oinarritua. (Egilea: Ziortza Guezuraga) Erresistentzia eskuratzen
4. irudia: Antibiotikoen ituak eta bakterioen erresistentzia nagusiak. Gerard D Wright-en irudian oinarritua. (Egilea: Ziortza Guezuraga) Erresistentzia eskuratzen Defentsa mekanismo horiek dira, hain zuzen, erresistentzia. Modu naturalean ematen den fenomenoa da, beraz. Naturala izanik ere, oso kontuan izateko bi faktore ditu erresistentziak: Antibiotikoen erabilera okerra eta erresistentzien zabalkuntza. Azken hau da bereziki kezkagarria, antibiotikoei aurre egiteko estrategiak bakterioz bakterio hedatzen dira, erresistentzia zabaltzen dela. Bi modu nagusitan gertatzen da erresistentziaren zabalkuntza:
Populazioaren biziraupena. Antibiotikoek hiltzen ez dituzten populazioak hazi egiten dira, bakterio erresistenteen biziraupena bermatzen duena.
Geneen transferentzia horizontala. Erresistente bilakatzen dituzten geneak bakterioz bakterio pasatzea. Hiru transferentzia mota daude:
1. Transformazioa. A bakterio batek DNA askatzen du mediora eta B Bakterioak DNA molekulak hartzen ditu mediotik, zuzenean hormatik zehar sartzen direnak.
2. Transdukzioa. Fagoak (birusak) baliatuta bakteria batetik bestera eramaten da DNA.
3. Konjugazioa. Bakterioen arteko kontaktu zuzenaren bidez egiten da transferentzia. Lehen aipatutako plasmidoak baliatuta geneak bakterio batetik bestera pasatzen dira.
Konjugazioa gertatzeko plasmido konjugatiboak behar dira, erresistentzia sortzen duten geneez gain, transferentzia aktibatzeko geneak ere dituztenak. Hiru pausutan ematen da:
- Bakterio emaileak pilusa sortzen du transferentzia geneei esker. Pilusa bi bakterioen arteko lotura izango da, pasabidea.
- Bi bakterioak elkartzean plasmidoa aldatu egiten da eta erlaxosoma bihurtzen da. Kate bikoitza dute plasmidoek eta horretako harizpi bat bakterio emailetik hartzailera pasatzen da.
- Bai bakterio emaileak bai hartzaileak harizpia kate bikoitz bihurtzen dute. Modu honetan, gene erresistenteak eta transferentzia geneak eskuratuta, emaile bilakatzen da ordura arte hartzailea izan den bakterioa.
 7. irudia: Konjugazioaren prozesua. (Egilea: Ziortza Guezuraga) Proteina motorraren bila
7. irudia: Konjugazioaren prozesua. (Egilea: Ziortza Guezuraga) Proteina motorraren bila Konjugazioa da, hain justu, Itziar Alkorta eta bere taldea ikertzen ari diren mekanismoa. Labur esanda, plasmidoa bakterio batetik bestera pasatzea ahalbidetzen duen proteina, T4CP izenekoa, ezeztatzea dute helburu. Akopladorea eta motor molekularra da T4CP proteina: ATP molekula hidrolizatzen du ADPa eta energia askatuz, eta askatzen den energia hori DNA harizpia beste bakteriora ponpatzeko erabiltzen du zelulak.
 8.irudia: “Superbakterioek” sortzen dituzten infekzioak osasun publikoko arazo larria bihurtzen ari dira, Bakterioen konjugazioa da antibiotikoen aurrean bakterioek azaleratzen duten barreiaduraren erantzulea. Prozesu horretan, batez ere, T4CP proteina aklopadorea da funtsezkoa. (Egilea: Ziortza Guezuraga)
8.irudia: “Superbakterioek” sortzen dituzten infekzioak osasun publikoko arazo larria bihurtzen ari dira, Bakterioen konjugazioa da antibiotikoen aurrean bakterioek azaleratzen duten barreiaduraren erantzulea. Prozesu horretan, batez ere, T4CP proteina aklopadorea da funtsezkoa. (Egilea: Ziortza Guezuraga) Konjugazioaren erdian dago proteina eta funtsezkoa da prozesua gerta dadin, T4CP proteinaren faltan bakterio konjugazioa ez da gertatzen. Mintz proteina da T4CP, ikertzeko zailak direnak, challenging proteins bezala ezagunak dira. Lan egiteko oso zailak dira, oso baldintza zehatzak behar dituzte eta datuak lortzea asko kostatzen da. 20 urte daramatzate proteina hau ikertzen.
Nahiz eta hasiera batean T4CP proteina baino ez zuten aztertzen, ikerketa taldeak hainbat proteina akopladore aztertzen ditu egun, ezaugarri komunak identifikatzeko. Batez ere, ATParen erabilerari dagokionez.
Teknika biokimiko, biofisiko eta genetikoak erabiltzen dituzte proteina hauek zelan funtzionatzen diren ikertzeko. Patroiak aurkitu eta inhibizioa lortzeko estrategia komuna garatu daiteke proteina akopladore guztietarako. Konjugazioa kontrolatu ahal izateko. Horretaz gain, alderdi aplikatua ere badu ikerketak.
- Kimiotekak aztertzen ari dira, bertako konposatuak aztertuta T4PC proteina inhibitzeko baliagarriak izan daitezkeenak aurkitzeko, hauen bidez bakterio batetik besterako transferentzia kontrolatzeko.
- Bilduma teorikoak ere erabiltzen ari dira. Kimiotekiko ezberdintasuna da inhibitzaile posibleak teorikoki aztertzen direla, proteinaren ezaugarriak kontuan izanda.
Bakterioen aurkako borrokan antibiotikoek egiten duten eraso zuzenarekin batera erresistentzien hedapenaren kontrola konbinatzea da ikerketaren helburua. Infekzioak kontrolatzeko antibiotikoak erabili behar dira, baina konjugazio inhibitzaileekin konbinatuta, erresistentzia eta erresistentziaren hedapena ekidin daitezke.
Bakterio infekziosoen aurkako borrokan, beraz, garrantzitsua da antibiotikoak topatu eta garatzeaz gain, bestelako estrategiekin konbinatzea. Sagua eta katua bezala, antibiotiko berriak sortzen zientziak eta erresistentziak garatzen bakterioak, ibili beharrean irtenbide definitiboagoak lortzeko.
Artikulu hau Itziar Alkortak Azkuna Zentroan emandako hitzaldian dago oinarritua:
———————————————————————–
Egileaz: Ziortza Guezuraga kazetaria eta UPV/EHUko Kultura Zientifikoko Katedraren kolaboratzailea da.
———————————————————————–
The post Antibiotikoak, bakterioak eta superbakterioak appeared first on Zientzia Kaiera.
El teorema de los cuatro colores (1): una historia que comienza en 1852
Wolfgang Haken
Smote the Kraken
One! Two! Three! Four!
Quoth he: “the monster is no more”.
(W.T. Tutte)
El teorema de los cuatro colores se enunció en forma de conjetura en 1852, pero tuvo que esperar hasta el año 1996 para ser admitido como válido por la práctica totalidad de la comunidad matemática.
El poema que abre este escrito es precisamente el que William Thomas Tutte dedicó a Wolfgang Haken –uno de los investigadores que dio una primera prueba del teorema en 1977, aunque tuvo que esperar hasta 1996 a ser ratificada por otros investigadores– tras más de cien años de fallidos intentos por probar su validez.
El teorema de los cuatro colores afirma que, para colorear un mapa geográfico plano sin que dos países colindantes tengan el mismo color, basta con cuatro tonos diferentes.
Se imponen las siguientes restricciones a los mapas aludidos en el teorema:
-
son siempre conexos –es decir, de una pieza– y cada una de sus regiones también es conexa;
-
dos territorios distintos de un mapa no pueden tocarse sólo en un punto, es decir, para ser colindantes deben compartir frontera en el sentido indicado en la figura 1.
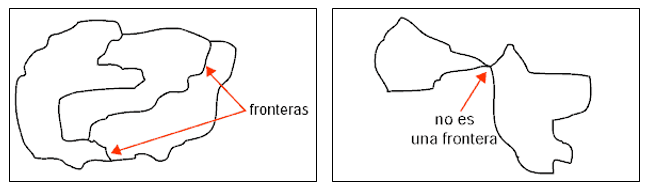
Figura 1.
Podría pensarse que un mapa con una gran cantidad regiones, poseyendo extrañas formas, y colindando de manera singular necesitaría mayor cantidad de colores que un mapa más sencillo. Pero, el teorema afirma que no es el caso.
Se trata de un problema topológico, ya que no es importante la forma de las regiones del mapa, sino la manera en la están colocadas las unas respecto a las otras.
¿Cómo empezó esta historia? El abogado y botánico Francis Guthrie observó que era capaz de colorear un mapa complicado de los cantones de Inglaterra con sólo cuatro colores, y conjeturó que debía suceder lo mismo con cualquier otro mapa geopolítico. En 1852, se lo comentó a su hermano Frederick y éste a su vez a Augustus de Morgan, su profesor.
Francis Guthrie observó, además, que tres colores no bastaban, mostrando lo que llamó el diagrama crítico (figura 2), que obviamente precisa de cuatro colores para no contradecir las condiciones de su conjetura –la parte blanca central es una región del mapa–.
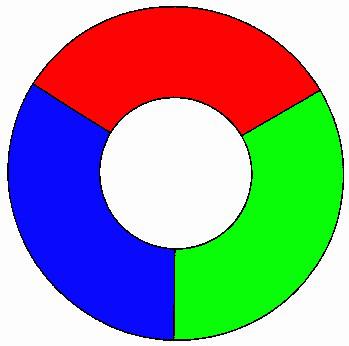
Figura 2: el diagrama crítico.
Frederick Guthrie fue el primero en observar que el problema de los cuatro colores no se podía generalizar a dimensión 3: en efecto, según un ejemplo posterior de Heinrich Tietze, es posible construir un ejemplo de mapa tridimensional que precise tantos colores como se desee. Su propuesta consistía en tomar barras numeradas de 1 hasta n,ordenándolas como muestra la figura 3, y sobre ellas colocaba otras n barras numeradas de 1 hasta n, girándolas 45 grados. De este modo, Tietze conseguía un mapa tridimensional conn regiones (cada número correspondía a un ‘país’), que obviamente necesitaba exactamente n colores para no contradecir las reglas de la conjetura…
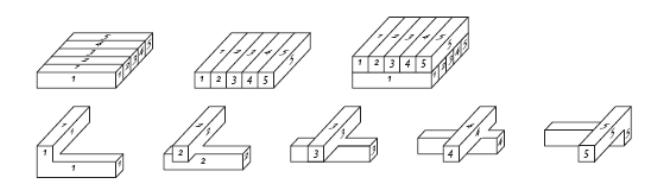
Figura 3: el ejemplo de Tietze para n=5.
Augustus de Morgan se interesó por la conjetura de Guthrie y la difundió entre sus colegas. El 23 de octubre de 1852 escribió a William Rowan Hamilton: “A student of mine [se refiere a Frederick Guthrie] asked me today to give him a reason for a fact which I did not know was a fact – and do not yet. He says that if a figure be any how divided and the compartments differently coloured so that figures with any portion of common boundary line are differently coloured – four colours may be wanted, but not more – the following is the case in which four colours are wanted. Query cannot a necessity for five or more be invented […]”

Figura 4: carta de De Morgan a Hamilton que incluye el diagrama crítico. Imagen Wikipedia.
En ese momento Hamilton estaba trabajando en teoría de cuaterniones, y respondió cuatro días después a De Morgan, con sarcasmo: “I am not going to attemt your quaternion of colour very soon”.
Decepcionado, De Morgan se puso en contacto con otros matemáticos. En 1853, escribió a William Whewell, describiendo la conjetura como un axioma matemático.
El problema de los cuatro colores cruzó el Atlántico: Charles Sanders Peirce dio un seminario sobre la demostración, aunque nunca la escribió.
Tras la muerte de De Morgan en 1871, el problema de los cuatro colores pareció olvidarse; aunque Peirce seguía buscando su demostración, ninguno de los amigos británicos de De Morgan lo mencionaban.
Años más tarde, en junio de 1878, Arthur Cayley hizo la siguiente pregunta en un encuentro de la London Mathematical Society: “Has a solution been given of the statement that in colouring a map of a country, divided into counties, only four colours are required, so that no two adjacent counties should be painted in the same colour”?.
Realmente interesado por el problema, en 1879 publicó una nota corta sobre el tema en los Proceedings of the Royal Geographical Society. Admitía la dificultad del tema: “I have not succeeded in obtaining a general proof: and it is worth while to explain wherein the difficulty consists”.
Entre otros, en esa nota observaba que, a la hora de probar el teorema, podían imponerse condiciones más restrictivas sobre los mapas a colorear, en particular, bastaba con limitarse a mapas cúbicos, es decir, aquellos en los que hay exactamente tres regiones en cada punto de encuentro. En efecto, si en un mapa hay más de tres regiones en alguno de los puntos de encuentro (ver figura 5); sobre este punto puede pegarse un pequeño parche que produce un mapa cúbico. Si se puede colorear este mapa con cuatro colores, también se puede obtener un 4-coloreado del mapa original: basta con colapsar el parche en un punto.
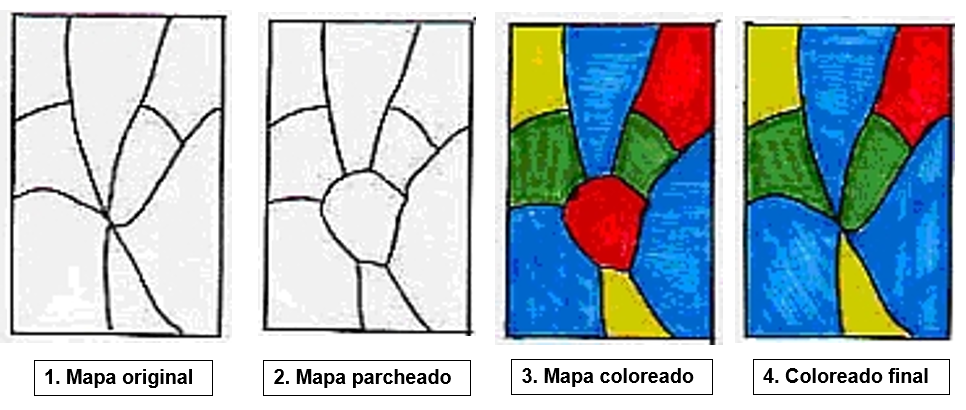
Figura 5 Basta con considerar mapas cúbicos.
Continuará en… El teorema de los cuatro colores (2): el error de Kempe y la clave de la prueba.
Referencias:
Marta Macho Stadler, Mapas, colores y números, Descubrir las matemáticas hoy: Sociedad, Ciencia, Tecnología y Matemáticas 2006 (2008) 41-68
Robin Wilson, Four colors suffice: how the map problem was solved, Princeton University Press, 2002.
Sobre la autora: Marta Macho Stadler es profesora de Topología en el Departamento de Matemáticas de la UPV/EHU, y colaboradora asidua en ZTFNews, el blog de la Facultad de Ciencia y Tecnología de esta universidad.
El artículo El teorema de los cuatro colores (1): una historia que comienza en 1852 se ha escrito en Cuaderno de Cultura Científica.
Entradas relacionadas:Bilbabio-izurria
 Juan Ignacio Pérez eta Miren Bego Urrutia Janaria
Juan Ignacio Pérez eta Miren Bego Urrutia Janaria ———————————————————————————————————–
Izurriteetan pentsatzen dugunean, intsektuak edo arratoiak izaten dira burura etorri ohi zaizkigun animaliak. Baina horiek ez dira izurritea sor dezaketen animalia bakarrak. Bibalbioen artean bada bat egiazko izurria sortu duena, oso arriskutsua gainera.
1. irudia: Zebra-muskuiluak. (Argazkia: D. Jude, Univ. of Michigan / Wikimedia Commons domeinu publikoko lana)
{kind=link}
2007ko abuztuan, ur gezetako bibalbio baten larbak aurkitu zituzten Gipuzkoako Lareoko urtegian; zebra-muskuiluaren larbez ari gara. Alarma piztu zuen aurkikuntzak, orduan ikusi baitziren lehenengoz larba horiek Kantauri Isuriko urtegi edo ibaietan. Ondorio larririk ez zuen ustekabeko gertaeratzat jo zen lehen agerpen hura, ondoren egindako ikerketen arabera ez baitzen larba horien presentzia baieztatu. Zoritxarrez, badago kezkatzeko arrazoirik, jakin denez Dreissena polymorpha bibalbioaren izurriak hedatzen jarraitu du-eta. Espezie inbaditzaile honen larbak Araban Zadorrako urtegi-sisteman 2011ko ekainean aurkitu ondoren, Bizkaian ere ale helduen presentzia baieztatu zen Arratia ibaiaren goiko aldeak dagoen Undurragako urtegian; heldu da zebra-muskuilua Kanturi Isurira beraz. 2011ko irailean jakin zen Urrunagako urtegiko erriberako beste zenbait aldetan ere atzeman zituztela. Tamaina txikikoak direnez aurkitutako bibalbio gehienak, populazioak finatzen hasiak baino ez daudela ondorioztatu zuten adituek, baina kontuan hartuta gure urtegi-sistema guztiak elkar lotuta daudela, litekeena da kolonia hauek hazten joatea eta nonahi ezartzea.
Zebra-muskuilua arriskutsua dela esan dugu; orain azalduko dugu zergatik. Azalpena eman aurretik, hala ere, espezieari buruzko zertzelada batzuk eskainiko ditugu. Dreissena polymorpha da zebra-muskuiluaren izen zientifikoa, eta paradoxikoa bada ere, ez da muskuilu bat; ezagutzen ditugun bibalbioen artean txirlak daude muskuiluak baino hurbilago zebra-muskuilutik. Muskuilu izena itxurari dagokio, muskuilu baten antza baitu, eta gainera, substratuan finkatzeko muskuiluek duten firuzko egitura bera dute, bisu izenekoa. Eta zebra esaten zaio maskor marraduna daukalako.

2. irudia: Zebra-muskuiluaren aleak aurkitu ziren Uztailaren 6ean Undurragako urtegian, Arratia ibaiaren arroan. (Argazkia: Araialdea Gaur)
Dreissena polymorpha ur gezetako bibalbio txikia da, luzeran 5 cm-ra irits daitekeena. Errusiako hego-ekialdeko aintziretan du jatorria, baina espezie inbaditzailea da, munduko toki askotatik sakabanatu baita. Gazitasun desberdineko uretan bizirik manten daiteke denbora luzez, eta tenperaturari dagokionez ere jasankortasun-tarte zabala du. Gainera, oso eraginkorra da: arrautza kopuru itzelak ekoizten ditu ur gezetako beste bibalbioekin erkatuz gero.
XIX. mendean hedatu zen Europako kanaletatik eta Ipar Itsasora heldu, bai eta Britainia Handira ere. Europako hegoaldea geroago kolonizatu zuen, Italiar eta Iberiar penintsuletara joan den mendearen azken laurdenera arte ez baitzen heldu. Ipar Amerikara 1988an iritsi zen, Laku Handietara, eta ordutik apurka-apurka Ipar Amerika osotik zabaltzen ari da. Aisialdirako belaontziak eta batelak ibai batetik bestera edo aintzira batetik bestera eramatean hedatu da horrenbeste, kroskoetan itsatsita bidaiatzen baitute muskuilu txikiek eta larbek. Horixe izan omen zen Lareoko urtegian gertatu zena. Hala ere, eta Ebro ibaia kolonizatu badu ere, Kantauri Isuriko ibaietara ez da oraindik heldu.
Beldurgarriak dira bibalbio honek sortzen dituen kalteak. Toki askotan, bertako bibalbio-espezieak guztiz ordezkatu ditu. Tokian betidanik egon diren bibalbio-populazioen gainetik finkatzeko gai dira; horrela, zebra-muskuiluaren populazio berriak sortu, eta azpian geratzen direnak hil egiten dira. Gainera, kaietan itsasten direnean kalteak sor ditzakete zurezko egituretan, bai eta industrietan hozte-prozesuetarako erabiltzen diren kanal, hodi eta ponpetan. Hodi batetik sartzen badira, urte gutxitan bete dezakete hodiaren argia guztiz itxi arte. Gastu handiak egin behar dira gero hodiak garbi mantentzeko.

3. irudia: Rutilus rutilus arraina zebra-muskuiluaz elikatzen da. (Argazkia: Cornelius Herget / Wikipedia CC BY-SA 3.0 lizentziapean)
Bestalde, bada zebra-muskuilua jaten duen arrainik: horietako bat Rutilus rutilus ur gezetako arraina da, eta, zenbait tokitan, zebra-muskuiluak dira arrain horrek jaten duenaren % 90. Hala ere, orain arte ez da inon ikusi arrainen harraparitzak Dreissenaren populazioak muga ditzakeenik.
Kontuak kontu, Dreissena polymorpha izurri arriskutsua dela esatea ez da inondik ere gehiegikeria bat; ibaietako zein aintziretako biodibertsitatearen arerio handia izateaz gain, izugarriak dira sor ditzakeen kalte ekonomikoak.
—————————————————–
Egileez: Juan Ignacio Pérez Iglesias (@Uhandrea) eta Miren Bego Urrutia Biologian doktoreak dira eta UPV/EHUko Animalien Fisiologiako irakasleak.
—————————————————–
Artikulua UPV/EHUren ZIO (Zientzia irakurle ororentzat) bildumako Animalien aferak liburutik jaso dugu.
The post Bilbabio-izurria appeared first on Zientzia Kaiera.
Bilbabio-izurria
 Juan Ignacio Pérez eta Miren Bego Urrutia Janaria
Juan Ignacio Pérez eta Miren Bego Urrutia Janaria ———————————————————————————————————–
Izurriteetan pentsatzen dugunean, intsektuak edo arratoiak izaten dira burura etorri ohi zaizkigun animaliak. Baina horiek ez dira izurritea sor dezaketen animalia bakarrak. Bibalbioen artean bada bat egiazko izurria sortu duena, oso arriskutsua gainera. 1. irudia: Zebra-muskuiluak. (Argazkia: D. Jude, Univ. of Michigan / Wikimedia Commons domeinu publikoko lana)
1. irudia: Zebra-muskuiluak. (Argazkia: D. Jude, Univ. of Michigan / Wikimedia Commons domeinu publikoko lana) 2007ko abuztuan, ur gezetako bibalbio baten larbak aurkitu zituzten Gipuzkoako Lareoko urtegian; zebra-muskuiluaren larbez ari gara. Alarma piztu zuen aurkikuntzak, orduan ikusi baitziren lehenengoz larba horiek Kantauri Isuriko urtegi edo ibaietan. Ondorio larririk ez zuen ustekabeko gertaeratzat jo zen lehen agerpen hura, ondoren egindako ikerketen arabera ez baitzen larba horien presentzia baieztatu. Zoritxarrez, badago kezkatzeko arrazoirik, jakin denez Dreissena polymorpha bibalbioaren izurriak hedatzen jarraitu du-eta. Espezie inbaditzaile honen larbak Araban Zadorrako urtegi-sisteman 2011ko ekainean aurkitu ondoren, Bizkaian ere ale helduen presentzia baieztatu zen Arratia ibaiaren goiko aldeak dagoen Undurragako urtegian; heldu da zebra-muskuilua Kanturi Isurira beraz. 2011ko irailean jakin zen Urrunagako urtegiko erriberako beste zenbait aldetan ere atzeman zituztela. Tamaina txikikoak direnez aurkitutako bibalbio gehienak, populazioak finatzen hasiak baino ez daudela ondorioztatu zuten adituek, baina kontuan hartuta gure urtegi-sistema guztiak elkar lotuta daudela, litekeena da kolonia hauek hazten joatea eta nonahi ezartzea.
Zebra-muskuilua arriskutsua dela esan dugu; orain azalduko dugu zergatik. Azalpena eman aurretik, hala ere, espezieari buruzko zertzelada batzuk eskainiko ditugu. Dreissena polymorpha da zebra-muskuiluaren izen zientifikoa, eta paradoxikoa bada ere, ez da muskuilu bat; ezagutzen ditugun bibalbioen artean txirlak daude muskuiluak baino hurbilago zebra-muskuilutik. Muskuilu izena itxurari dagokio, muskuilu baten antza baitu, eta gainera, substratuan finkatzeko muskuiluek duten firuzko egitura bera dute, bisu izenekoa. Eta zebra esaten zaio maskor marraduna daukalako.
 2. irudia: Zebra-muskuiluaren aleak aurkitu ziren Uztailaren 6ean Undurragako urtegian, Arratia ibaiaren arroan. (Argazkia: Araialdea Gaur)
2. irudia: Zebra-muskuiluaren aleak aurkitu ziren Uztailaren 6ean Undurragako urtegian, Arratia ibaiaren arroan. (Argazkia: Araialdea Gaur) Dreissena polymorpha ur gezetako bibalbio txikia da, luzeran 5 cm-ra irits daitekeena. Errusiako hego-ekialdeko aintziretan du jatorria, baina espezie inbaditzailea da, munduko toki askotatik sakabanatu baita. Gazitasun desberdineko uretan bizirik manten daiteke denbora luzez, eta tenperaturari dagokionez ere jasankortasun-tarte zabala du. Gainera, oso eraginkorra da: arrautza kopuru itzelak ekoizten ditu ur gezetako beste bibalbioekin erkatuz gero.
XIX. mendean hedatu zen Europako kanaletatik eta Ipar Itsasora heldu, bai eta Britainia Handira ere. Europako hegoaldea geroago kolonizatu zuen, Italiar eta Iberiar penintsuletara joan den mendearen azken laurdenera arte ez baitzen heldu. Ipar Amerikara 1988an iritsi zen, Laku Handietara, eta ordutik apurka-apurka Ipar Amerika osotik zabaltzen ari da. Aisialdirako belaontziak eta batelak ibai batetik bestera edo aintzira batetik bestera eramatean hedatu da horrenbeste, kroskoetan itsatsita bidaiatzen baitute muskuilu txikiek eta larbek. Horixe izan omen zen Lareoko urtegian gertatu zena. Hala ere, eta Ebro ibaia kolonizatu badu ere, Kantauri Isuriko ibaietara ez da oraindik heldu.
Beldurgarriak dira bibalbio honek sortzen dituen kalteak. Toki askotan, bertako bibalbio-espezieak guztiz ordezkatu ditu. Tokian betidanik egon diren bibalbio-populazioen gainetik finkatzeko gai dira; horrela, zebra-muskuiluaren populazio berriak sortu, eta azpian geratzen direnak hil egiten dira. Gainera, kaietan itsasten direnean kalteak sor ditzakete zurezko egituretan, bai eta industrietan hozte-prozesuetarako erabiltzen diren kanal, hodi eta ponpetan. Hodi batetik sartzen badira, urte gutxitan bete dezakete hodiaren argia guztiz itxi arte. Gastu handiak egin behar dira gero hodiak garbi mantentzeko.
 3. irudia: Rutilus rutilus arraina zebra-muskuiluaz elikatzen da. (Argazkia: Cornelius Herget / Wikipedia CC BY-SA 3.0 lizentziapean)
3. irudia: Rutilus rutilus arraina zebra-muskuiluaz elikatzen da. (Argazkia: Cornelius Herget / Wikipedia CC BY-SA 3.0 lizentziapean) Bestalde, bada zebra-muskuilua jaten duen arrainik: horietako bat Rutilus rutilus ur gezetako arraina da, eta, zenbait tokitan, zebra-muskuiluak dira arrain horrek jaten duenaren % 90. Hala ere, orain arte ez da inon ikusi arrainen harraparitzak Dreissenaren populazioak muga ditzakeenik.
Kontuak kontu, Dreissena polymorpha izurri arriskutsua dela esatea ez da inondik ere gehiegikeria bat; ibaietako zein aintziretako biodibertsitatearen arerio handia izateaz gain, izugarriak dira sor ditzakeen kalte ekonomikoak.
—————————————————–
Egileez: Juan Ignacio Pérez Iglesias (@Uhandrea) eta Miren Bego Urrutia Biologian doktoreak dira eta UPV/EHUko Animalien Fisiologiako irakasleak.
—————————————————–
Artikulua UPV/EHUren ZIO (Zientzia irakurle ororentzat) bildumako Animalien aferak liburutik jaso dugu.
The post Bilbabio-izurria appeared first on Zientzia Kaiera.
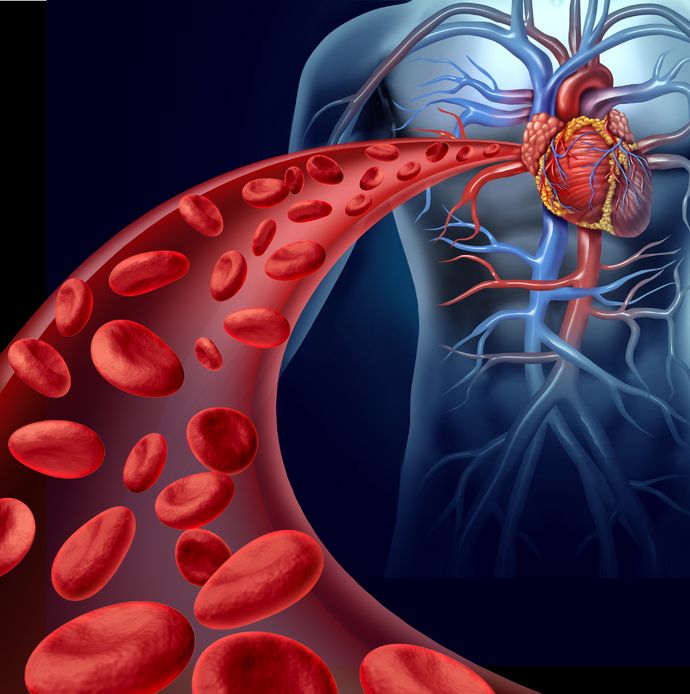
El flujo sanguíneo se reorganiza en respuesta a las necesidades
Como vimos aquí, en reposo, el corazón humano bombea sangre a razón de 5 litros por minuto (l m-1). La sangre que sale por la aorta y se dirige a los tejidos procedente de los pulmones va cargada de O2. Veamos cómo se reparte ese flujo entre unos órganos y otros.
El sistema digestivo recibe 1,35 l min-1, un 27% del total. El tracto digestivo recibe 1 l min-1, y el resto 0,35 l min-1, aproximadamente, va directamente al hígado, aunque este órgano recibe, a través del sistema portal hepático, un flujo adicional procedente precisamente del intestino con sustancias recién absorbidas. Este flujo sanguíneo proporciona oxígeno a un tejido metabólicamente muy activo. Pero también está al servicio de la incorporación al organismo de los nutrientes digeridos y absorbidos, y su posterior remisión al hígado para su posible almacenamiento o transformación, y al resto de los tejidos para su uso; en este segundo caso, no obstante, la sangre ha de volver al corazón para su redistribución posterior.
El segundo destino que más sangre recibe en reposo son los riñones, a los que se dirige un 20% del total de sangre bombeada en un ciclo de contracción-relajación del ventrículo izquierdo del corazón. El flujo que reciben los riñones es de 1 l min-1. Aunque las células renales son muy activas metabólicamente por el trabajo de transporte iónico que efectúan, la principal razón de ese elevado flujo es la necesidad de filtrar la sangre en los glomérulos renales para limpiarla de las sustancias de deshecho (urea, principalmente) y para ajustar los elementos del balance de agua y sales, de manera que tal balance resulte equilibrado y pueda mantenerse la constancia en las propiedades osmóticas e iónicas del medio interno.
La muscultura esquelética recibe 750 ml min-1 (un 15%) del flujo sanguíneo. Puede parecer una cifra importante, pero no lo es en términos relativos, ya que la musculatura esquelética representa alrededor de la mitad de la masa corporal.
Por comparación, mayor importancia relativa tiene el flujo de sangre que riega el encéfalo: 650 ml min-1(13%). No debe sorprender ese valor, dado que el tejido encefálico es el de mayor actividad metabólica: algo más del 20% de la del conjunto del organismo en reposo. Ese elevado metabolismo es el que alimenta energéticamente el mantenimiento de los gradientes iónicos de los que depende la transmisión de los impulsos nerviosos, así como el conjunto de procesos implicados en las sinapsis químicas. Hay que tener en cuenta que la masa encefálica es de entre 1,3 y 1,5 kg. dependiendo de la talla del individuo, o sea, representa del orden del 2% de la masa corporal solamente.
La piel recibe 450 ml min-1 (un 9%), el corazón 150 ml min-1 (3%) y los huesos y resto de elementos corporales, 650 ml min-1 (13%).
Las cosas cambian de forma sustancial si el individuo debe hacer alguna actividad física. Para empezar, en condiciones de ejercicio moderado el gasto cardiaco se multiplica por 2,5: pasa a ser de 12,5 l min-1. Pero el cambio no consiste solo en un aumento del flujo sanguíneo total; también cambia, y mucho, el reparto de ese flujo entre los diferentes órganos.
El cambio más importante es el que registra el riego muscular. Se multiplica por más de diez y de los 750 ml min-1 en reposo pasa a 8 l min-1; o sea, casi dos terceras partes (64%) del flujo total se dirige a la musculatura esquelética bajo esas condiciones. La razón de ese aumento no requiere explicaciones adicionales. La piel y el corazón son los otros órganos cuya irrigación aumenta de forma importante. La piel pasa a recibir 1,7 l min-1 (el 13,6%); no hay que perder de vista que la piel es el principal enclave a través del cual disipamos energía en forma de calor hacia el exterior. Y el aumento de la irrigación cardiaca, que pasa a ser de 550 ml min-1 (4,4 %) obedece, precisamente, a la necesidad de aumentar el gasto cardiaco elevando la frecuencia de latido.
Nada sorprendentemente, el encéfalo recibe el mismo riego sanguíneo (650 ml min-1) que en reposo, lo que da idea de la importancia de mantener la actividad encefálica constante, incluso cuando otras requieren aportes adicionales de O2 y nutrientes.
Y luego están los órganos cuyo riego disminuye, tanto en términos absolutos como relativos, porque las funciones que desempeñan permiten una reducción a veces importante, ya que no es esencial que se mantengan a un nivel constante en todo momento. El tracto digestivo pasa a recibir 600 ml min-1 (4,8%), los riñones, 550 ml min-1 (4,4%) y huesos y demás elementos, 450 ml min-1 (3,6%).
Los ajustes necesarios para redirigir el flujo sanguíneo se producen en el sistema de arteriolas. Se trata del sistema que, en conjunto, más resistencia ofrece a la circulación de la sangre; por esa razón es en esa parte de la circulación general donde se produce la caída más grande de la presión sanguínea: su valor medio pasa de unos 90 mm Hg al salir de las grandes arterias, a los aproximadamente 40 mm Hg al llegar a los capilares. Pues bien, pequeñas modificaciones en el tono arteriolar, aumentando o disminuyendo el diámetro interno de los vasos de unas zonas o de otras, dan lugar a cambios muy grandes en el flujo de sangre a través de los diferentes tejidos. Y esos cambios responden a señales intrínsecas. Esto no implica que no haya control extrínseco del diámetro de las arteriolas. Lo hay y corre a cargo del sistema nervioso simpático, pero, al parecer, el objeto de tal control no es el de regular el flujo, sino controlar la presión arterial en su conjunto.
Las señales que generan los cambios locales de flujo son químicas y tienen que ver con la actividad metabólica de los tejidos afectados. Se trata, por ello, de un control local, intrínseco. La reducción de la concentración de O2 en la sangre produce vasodilatación arteriolar. Así, cuando el consumo de oxígeno es alto en los músculos esqueléticos, su concentración baja rápidamente y ese descenso provoca una mayor apertura de las arteriolas que riegan esos músculos. Además de la disminución de la concentración de O2, el aumento de la de CO2 –provocada por la misma causa- tiene el mismo efecto. La acidificación del entorno –por efecto de la combinación de CO2 con el agua y por el aumento en la concentración de lactato- da lugar también a un aumento en el diámetro de las arteriolas. Otro factor es la elevación del K+ extracelular; esa elevación se produce cuando las bombas Na+-K+ no son capaces de reintroducir en las células (neuronas encefálicas o células musculares) el K+ que sale al exterior como consecuencia de los frecuentes potenciales de acción que ocurren en zonas especialmente activas. Una concentración osmótica elevada tiene el mismo efecto; ocurre cuando el metabolismo celular es muy alto y da lugar a la producción excesiva de moléculas osmóticamente activas, algunas de las cuales abandonan las células hacia el líquido intersticial. Y en el músculo cardiaco especialmente, la liberación de adenosina que se produce en respuesta a una elevada actividad metabólica o falta de oxígeno, también provoca un aumento del diámetro arteriolar y, por lo tanto, del flujo sanguíneo hacia la zona afectada.
Cuando, por las razones que sea, eches a correr y experimentes un aumento del ritmo cardiaco y un mayor flujo de calor hacia la superficie de tu cuerpo, quizás recuerdes esto que has leído aquí.
El artículo El flujo sanguíneo se reorganiza en respuesta a las necesidades se ha escrito en Cuaderno de Cultura Científica.
Entradas relacionadas:Los cimientos de la química neumática al estilo van Helmont (y 3)
El principio de conservación de la materia es algo que está implícito en van Helmont ya que asume en sus experimentos que se conserva independientemente de los cambios que sufra. Extendió esta idea a los metales, algo que los alquimistas veían de forma completamente opuesta. Según van Helmont, cuando los metales se disolvían en los ácidos no se destruían sino que se podían recuperar gramo por gramo (cosa muy útil si te persiguen los nazis).

Angelo Sala. Imagen: Wellcome Library, London. Wellcome Images
Estas ideas, sin embargo, no eran exclusivas de van Helmont. Por ejemplo, su contemporáneo Angelo Sala describe en Anatomia vitrioli (1617) como había disuelto una cierta cantidad de cobre en ácido sulfúrico, recuperado químicamente el cobre metálico y encontrado que pesaba exactamente lo mismo que el cobre con el que había comenzado el experimento.
No solo eso, Sala también demostró algo muy importante y que hoy se da (o debería darse) por sentado: pudo preparar en el laboratorio un sulfato de cobre hidratado que era exactamente igual al sulfato de cobre hidratado que se encuentra en la naturaleza, una idea revolucionaria para la época. Pero Sala no tenía la influencia y el prestigio de van Helmont, por lo que hoy no aparece en casi ningún libro de historia de la química, entre otras cosas, porque sus ideas más que notables, no tuvieron impacto.
Sin embargo, van Helmont tenía tanta influencia que incluso sus especulaciones más evidentemente arbitrarias tuvieron eco. Así, van Helmont postuló la existencia del alcahest, el disolvente universal, que tenía la propiedad de devolver las sustancias a sus estado primitivo, esto es, a agua, el componente básico de todo. Los químicos contemporáneos, que habían dejado de buscar la piedra filosofal, emplearon mucho tiempo y esfuerzo buscando el alcahest, incluido el famoso Robert Boyle.

La química moderna empezará realmente con la investigación de los gases por los llamados químicos neumáticos y será van Helmont el que establezca las bases de esta nueva área de investigación. Eso sí, con la esquizofrenia característica del siglo.
Para van Helmont como el aire no podía convertirse en agua, tenía que ser un elemento diferente. Sin embargo, existía una serie de sustancias “parecidas al aire” que aparecían frecuentemente en el transcurso de las reacciones químicas. Para referirse a ellas van Helmont empezó a utilizar el término gas, que en neerlandés suena muy parecido a caos, un término que Paracelso usó para referirse al aire en otro sentido.
Obviamente, para van Helmont los gases eran simplemente agua, no aire, ya que cualquier sustancia se convertía en gas e iba a la atmósfera por la muerte de sus “fermentos”. Un gas era caos porque no tenía forma. Un gas podía condensar en vapor (visible como distinto del aire) y caer en forma de lluvia (precipitar) por la influencia del blas, un término que la jerga química no terminó de aceptar y con el que van Helmont se refería a una especie de influencia o poder astral, “gravitacional”, que producía el cambio y el movimiento en el universo.
Como hemos visto, el siglo XVII produce grandes avances en la química pero también modelos teóricos arcaicos o arcaizantes. Serán ya otros nombres, todavía arrastrando algunas ideas alquímicas, los que comiencen este mismo siglo con la historia moderna de la química: Robert Boyle, Robert Hooke, John Mayow, John Joachim Becher, Georg Ernest Stahl o Stephen Hales ya no son alquimistas.
Sobre el autor: César Tomé López es divulgador científico y editor de Mapping Ignorance
El artículo Los cimientos de la química neumática al estilo van Helmont (y 3) se ha escrito en Cuaderno de Cultura Científica.
Entradas relacionadas:- Las transmutaciones de van Helmont (2)
- Jan van Helmont, filósofo por el fuego (1)
- El siglo de la esquizofrenia química
Wolframa, metal baliotsua oraindik ere!

1. irudia: Elhuyar anaiek, wolframa isolatu zuten 783ko irailaren 28an eta “wolfram” izena eman zioten sortu zen materia gogoan izateko.
Wolframa isolatzea Euskalerriaren Adiskideen Elkarteak 7 urte lehenago sortutako “Real Seminario Patriotico Bascongado”aren lorpen zientifiko ikusgarria izan zen, eta agian Euskal Herriak inoiz zientzia arloan egin duen eta egingo duen aurkikuntzarik handienetakoa. Baina proiektua bultzatu zutenek zientziaren oso ikuspegi utilitarista zeukaten eta beraien proiektuari ospe handia eman dion lorpena izan arren, badirudi metal berriari ez ziotela inongo aplikaziorik aurkitu. Aplikazioak oso poliki joan ziren iristen, hurrengo mendean hasita, baina iritsi dira. Eta wolframa elementu estrategikoa izatera iritsi da, gaurko gizartearen garapenerako funtsezko elementuetako bat dela esan dezakegu, bere aplikazioei erreparatzen badiogu.
Ezaugarri bereziak ditu, aplikagarritasunari begira eragin handia dutenak: metal oso gogorra da, xaflakorra eta fusio-punturik altuena duen metala da (3683ºk/3410ºC). Wolframaren aplikazio batzuk aski ezagunak dira. Beroaren aurrean duen erresistentzia ezinbestekoa izan da gaur egun arte etxeetan erabili ditugun bonbillen funtzionamendurako. XX. mendearen hasieran Coolidge jaunak wolframezko filamentu harikorra txertatu zuen lehen aldiz bonbilletan eta horrek argiztapen-sistema elektrikoaren historian aurrerapen nabarmena eragin zuen. Bigarren Mundu Gerran garrantzi handia izan zuen metal horrek, blindajeetan erabiltzen delako; eta ondoren soldatze lanetara, makina-erreminta arlora eta milaka aplikazio desberdinetara zabaldu da erabilera. Laboratorium museoak dohaintzan jasotako wolframezko lehen pieza, “hegazkin motor bateko pieza; CFM56 modelokoa (wolframezko aleazioa)”, aplikazio industrial anitzen adibidea da.
Baina aldi berean, esparru oso berrietan ere bere izaera estrategikoa ari da erakusten. Horren lekukoa da bigarren dohaintza, “Wolfram purua (barra)”, ESS Bilbaok dohaintzan emandakoa.

2. Wolframita. (Argazkia: Laboratorium Museoa)
Europako Espalazio bidezko Neutroi Iturria (European Spallation Source-ESS) Europako egitasmo zientifiko garrantzitsuenetako bat da. Instalazio nagusia Suediako Lund hirian ari dira eraikitzen, baina proiektuak Zamudion ere badu bere bigarren egoitza (ESS Bilbao). ESS proiektuak oinarrizko zientziaren zein teknologiaren hainbat ikerketei bide berriak zabalduko dizkiola aurreikusten da.
Espalazioa neutroiak kopuru handian sortzeko aukera ematen duen teknika da, protoietatik abiatuta. Espalazioak egiten duena da atomo baten nukleoa ezpaldu, nukleoak neutroi-pultsuak emititu ditzan. Prozesu hori sortzeko, beharrezkoa da protoi multzo bat eremu elektromagnetikoen bidez azeleratzea, energia kantitate handia eskuratzen duten arte. Une horretan, material astun baten kontra zuzentzen dira, eta talkak abiatzen du neutroiak sortzen duen erreakzio nuklearra. Material astun hori, noski, wolframa da.
Espalazioa egiteko ingeniaritza-gailuak, wolframezko 36 sektore ditu (elementu horrekin egindako 7.000 pieza guztira), 2,5 metroko diametroa duen gurpil baten barruan kokatuta. Gurpilak 30 rpm-ko abiaduran biratuko du eta horrela protoi-sortak sektore bati eragingo dio aldiko, neutroiak lortuz. Zirkuitu osoa helio bidez hozten da. Guk dohaintzan jaso duguna 7.000 pieza horietako bat da.
Wolframa fusio nuklearretako instalakuntza esperimentaletan ere erabiltzen da, dibertoreetan hain zuzen ere, hau da, karga termiko handiena jasaten duten sistemetan. Aplikazio berri hau ezagutzera emateko, Erresuma Batuko Culham Centre for Fusion Energy (CCFE) erakundeak maileguan utzitako dibertoreko pieza bat izan genuen museoan ikusgai, aldi baterako.
—————————————————–
Egileez: Bergarako Laboratorium Museoko talde teknikoa.
Errekalde Jauregia, Juan Irazabal pasealekua, 1. 20570 Bergara
Harremanetarako: 943 769 003 eta laboratorium@bergara.eus
—————————————————–
![]()
The post Wolframa, metal baliotsua oraindik ere! appeared first on Zientzia Kaiera.
Wolframa, metal baliotsua oraindik ere!
 1. irudia: Elhuyar anaiek, wolframa isolatu zuten 783ko irailaren 28an eta “wolfram” izena eman zioten sortu zen materia gogoan izateko.
1. irudia: Elhuyar anaiek, wolframa isolatu zuten 783ko irailaren 28an eta “wolfram” izena eman zioten sortu zen materia gogoan izateko. Wolframa isolatzea Euskalerriaren Adiskideen Elkarteak 7 urte lehenago sortutako “Real Seminario Patriotico Bascongado”aren lorpen zientifiko ikusgarria izan zen, eta agian Euskal Herriak inoiz zientzia arloan egin duen eta egingo duen aurkikuntzarik handienetakoa. Baina proiektua bultzatu zutenek zientziaren oso ikuspegi utilitarista zeukaten eta beraien proiektuari ospe handia eman dion lorpena izan arren, badirudi metal berriari ez ziotela inongo aplikaziorik aurkitu. Aplikazioak oso poliki joan ziren iristen, hurrengo mendean hasita, baina iritsi dira. Eta wolframa elementu estrategikoa izatera iritsi da, gaurko gizartearen garapenerako funtsezko elementuetako bat dela esan dezakegu, bere aplikazioei erreparatzen badiogu.
Ezaugarri bereziak ditu, aplikagarritasunari begira eragin handia dutenak: metal oso gogorra da, xaflakorra eta fusio-punturik altuena duen metala da (3683ºk/3410ºC). Wolframaren aplikazio batzuk aski ezagunak dira. Beroaren aurrean duen erresistentzia ezinbestekoa izan da gaur egun arte etxeetan erabili ditugun bonbillen funtzionamendurako. XX. mendearen hasieran Coolidge jaunak wolframezko filamentu harikorra txertatu zuen lehen aldiz bonbilletan eta horrek argiztapen-sistema elektrikoaren historian aurrerapen nabarmena eragin zuen. Bigarren Mundu Gerran garrantzi handia izan zuen metal horrek, blindajeetan erabiltzen delako; eta ondoren soldatze lanetara, makina-erreminta arlora eta milaka aplikazio desberdinetara zabaldu da erabilera. Laboratorium museoak dohaintzan jasotako wolframezko lehen pieza, “hegazkin motor bateko pieza; CFM56 modelokoa (wolframezko aleazioa)”, aplikazio industrial anitzen adibidea da.
Baina aldi berean, esparru oso berrietan ere bere izaera estrategikoa ari da erakusten. Horren lekukoa da bigarren dohaintza, “Wolfram purua (barra)”, ESS Bilbaok dohaintzan emandakoa.
 2. Wolframita. (Argazkia: Laboratorium Museoa)
2. Wolframita. (Argazkia: Laboratorium Museoa) Europako Espalazio bidezko Neutroi Iturria (European Spallation Source-ESS) Europako egitasmo zientifiko garrantzitsuenetako bat da. Instalazio nagusia Suediako Lund hirian ari dira eraikitzen, baina proiektuak Zamudion ere badu bere bigarren egoitza (ESS Bilbao). ESS proiektuak oinarrizko zientziaren zein teknologiaren hainbat ikerketei bide berriak zabalduko dizkiola aurreikusten da.
Espalazioa neutroiak kopuru handian sortzeko aukera ematen duen teknika da, protoietatik abiatuta. Espalazioak egiten duena da atomo baten nukleoa ezpaldu, nukleoak neutroi-pultsuak emititu ditzan. Prozesu hori sortzeko, beharrezkoa da protoi multzo bat eremu elektromagnetikoen bidez azeleratzea, energia kantitate handia eskuratzen duten arte. Une horretan, material astun baten kontra zuzentzen dira, eta talkak abiatzen du neutroiak sortzen duen erreakzio nuklearra. Material astun hori, noski, wolframa da.
Espalazioa egiteko ingeniaritza-gailuak, wolframezko 36 sektore ditu (elementu horrekin egindako 7.000 pieza guztira), 2,5 metroko diametroa duen gurpil baten barruan kokatuta. Gurpilak 30 rpm-ko abiaduran biratuko du eta horrela protoi-sortak sektore bati eragingo dio aldiko, neutroiak lortuz. Zirkuitu osoa helio bidez hozten da. Guk dohaintzan jaso duguna 7.000 pieza horietako bat da.
Wolframa fusio nuklearretako instalakuntza esperimentaletan ere erabiltzen da, dibertoreetan hain zuzen ere, hau da, karga termiko handiena jasaten duten sistemetan. Aplikazio berri hau ezagutzera emateko, Erresuma Batuko Culham Centre for Fusion Energy (CCFE) erakundeak maileguan utzitako dibertoreko pieza bat izan genuen museoan ikusgai, aldi baterako.
—————————————————–
Egileez: Bergarako Laboratorium Museoko talde teknikoa.
Errekalde Jauregia, Juan Irazabal pasealekua, 1. 20570 Bergara
Harremanetarako: 943 769 003 eta laboratorium@bergara.eus
—————————————————–
![]()
The post Wolframa, metal baliotsua oraindik ere! appeared first on Zientzia Kaiera.
Historias de la malaria: El parásito
Terciana: Calentura intermitente que repite cada tercer día.
Cuartana: Calentura, casi siempre de origen palúdico, que entra con frío, de cuatro en cuatro días.
(Diccionario de la Lengua Española).
Fue el médico militar francés Alphonse Laveran, del que hablaré en otro capítulo de esta serie, quien demostró en 1880, con la ayuda de un microscopio no muy potente, que el parásito que provoca la malaria tenía, según vio en preparaciones de sangre fresca de enfermos, un cuerpo globular ciliado que, años más tarde se asignó a un protozoo del género Plasmodium. Ahora nosotros vamos a conocer en detalle al protagonista principal de las historias de la malaria.

Plasmodium falciparum
Son cuatro las especies de Plasmodium que causan la malaria en nuestra especie. El Plasmodium falciparum es la especie más letal, la más asociada a la mortalidad que provoca el parásito de la malaria. Menos letales son las otras tres especies implicadas: Plasmodium vivax, Plasmodium malariae y Plasmodium ovale.
El ciclo vital de los plasmodios tiene dos fases: una sexual en la hembra del mosquito transmisor, y otra asexual en el huésped vertebrado. Esta fase en el vertebrado tiene, a su vez, dos fases: la primera, una vez llega por la picadura del mosquito, en las células del hígado, y la segunda en los glóbulos rojos de la sangre. La descarga de los parásitos en gran cantidad desde los glóbulos rojos es la que provoca las fiebres típicas de la malaria.
Una vez que el insecto pica al vertebrado huésped, y se carga de células sexuales del parásito y, en el estómago del insecto, se unen, forman el zigoto o huevo fecundado que, a su vez, se rompe en abundantes células del protozoo llamadas esporocitos que llegan a las glándulas salivares del mosquito y, cuando éste pica, son descargados a la sangre del huésped vertebrado, y el ciclo comienza de nuevo.
Los esporocitos atraviesan la pared del digestivo del vertebrado y llegan al hígado, en una hora más o menos, y allí se multiplican formando los merozoitos que se descargan a la sangre e invaden los glóbulos rojos. Se vuelven a dividir hasta formar más merozoitos y, finalmente, las células sexuales que, cuando pica el mosquito, llegan a su estómago, y vuelve a comenzar el ciclo.
Complicado pero así consiguen tantos parásitos el éxito evolutivo que tienen.
La reproducción del Plasmodium en los glóbulos rojos tarda unas 48 horas en el falciparum, vivax y ovale, y 72 horas en malariae. El ciclo de 48 horas es el que provoca las fiebres que se llaman tercianas, con un acceso un día de cada dos. Si es con falciparum, es la terciana maligna; si es con ovale o vivax, es la fiebre terciana benigna. En malariae, con el ciclo de 72 horas, aparecen la fiebre que se denomina cuartana, con un día de cada tres.
El ciclo completo del desarrollo de los protozoos en la especie humana es de 9 a 15 días en falciparum, de 12 días en vivax y ovale, y de 20 días en malariae. Los parásitos no sobreviven en el huésped más de un año, para el falciparum, o de dos años, para el vivax y el ovale. El P. malariae plantea más problemas pues puede mantenerse hasta algunas decenas de años. En conclusión, si se controlan los insectos durante tres años en un determinado enclave con malaria, el parásito desaparece. Se hizo en la década de los sesenta en Estados Unidos, Puerto Rico y otros países y se erradicó la enfermedad.
La entrada de los protozoos en los glóbulos rojos se consigue por la unión del parásito a receptores de membrana de las células de la sangre. Si no hay unión, el plasmodio no puede entrar en el glóbulo rojo y completar su ciclo de crecimiento y, por tanto, no hay ataques de fiebre en el enfermo. Es lo que ocurre en África con el vivax que no consigue entrar en los glóbulos rojos. Es una especie que no aparece en muchas zonas de África, precisamente las mismas zonas en que gran parte de la población, entre el 90% y el 100%, tiene la mutación negativa del grupo sanguíneo Duffy. En esta mutación, el receptor de membrana, una proteína que caracteriza al grupo Duffy, no se sintetiza y, por tanto, no está en la membrana de los glóbulos rojos. Y el vivax, para entrar en la célula, se une precisamente a ese receptor Duffy. Y, por ello, al faltar el receptor no puede entrar en el glóbulo rojo. Se ha propuesto que por esta causa ha desaparecido de grandes zonas de África. Está en chimpancés y gorilas, pero no en la especie humana.
El ciclo vital del parásito en el mosquito necesita de una temperatura ambiente elevada. En el Plasmodium falciparum es de más de 26ºC, con una mínima para poder desarrollarse de 18ºC, y el ciclo dura diez días. En vivax es de 8-9 días, en ovale son 12-14 días, y en malariae es de 14-15 días. Estas especies están, por esa restricción de temperaturas, confinadas al trópico y, si aparecen en zonas más templadas, es porque el mosquito vive en interiores, climatizados por nuestra especie, con un microclima más estable y temperaturas no tan bajas como en el exterior.
Con estas limitaciones de temperatura, es indudable el impacto que puede tener el calentamiento global para un aumento del área de distribución del Plasmodium. Ya está ocurriendo en el sur y el este de África, y la epidemia de malaria en Madagascar a finales de los ochenta, a partir de 1987, en zonas de altitud y con clima más fresco que en la costa, donde la enfermedad, con pequeños focos, había desaparecido en 1960, es un buen ejemplo. En pocos años alcanzó los 25000 muertos.
El origen y la expansión del plasmodio es anterior a la especie humana. La propagación por todo el planeta tuvo, cuando ya parasitaba a nuestra especie, precisamente en la extraordinaria movilidad y adaptación a todos los entornos del Homo sapiens.
Su origen según algunos expertos, está en algún protozoo, parásito del intestino de vertebrados, que consiguió atravesar la pared del tubo digestivo y adaptarse a vivir en otros órganos y, muy importante, en la sangre del huésped. Después, en un segundo paso, encontró un huésped invertebrado en los mosquitos picadores y chupadores de sangre. Hay que recordar que mosquitos parecidos a los actuales ya existían hace unos 150 millones de años.
Las líneas más antiguas de plasmodios de la malaria están en reptiles y aves y, en una línea más reciente, aparece en mamíferos, incluyendo a primates y a nuestra especie. Algunos dicen que el paso de falciparum de aves, quizá domésticas, a Homo sapiens, es reciente, de hace entre 5000 y 10000 años. Otros autores proponen que el parásito de humanos viene del chimpancé y que el paso se puede fechar hace unos 8 millones de años, cuando aparecieron las especies que llevarían, por una parte, a los chimpancés actuales y, por otra, a la especie humana. Las pruebas directas más antiguas de la malaria en la especie humana, aún con descripciones médicas y literarias anteriores, están en las momias egipcias, desde hace unos 4000 años, y en los cadáveres de cementerios cercanos a Roma y fechados hace unos 2000 años, en la época imperial. El análisis de ADN y pruebas inmunológicas de esos cuerpos han demostrado la presencia de falciparum. Incluso la momia de Tutankamon y las de tres familiares suyos, enterrados hace unos 3500 años, dan positivo al análisis de ADN de falciparum.
Ahora empezamos a conocer el genoma de los plasmodios. Tienen 14 cromosomas. Y el estudio del ADN lleva a estudiar su evolución. Por ejemplo, algunos de los plasmodios de humanos, en los análisis genéticos, son muy parecidos a los que se encuentran en otros monos actuales y, por tanto, el paso de la malaria a humanos sería más reciente. Así, el protozoo más cercano al falciparum se encuentra en los gorilas, y el de vivax en gorilas y chimpancés.
Además, el análisis genético ha permitido detectar que hay infecciones mixtas, con más de una de las especies de plasmodios en las zonas donde se localiza más de una. Es interesante que las infecciones con vivax o con malariae reducen la gravedad de la infección con falciparum. Incluso se ha encontrado otro plasmodio, parásito del macaco, que, a veces, también parásita a la especie humana. Es el Plasmodium knowlesi.
En cuanto al origen geográfico de la malaria en nuestra especie parece que se localiza en el centro y el este de África, en la región de Etiopía. Desde allí, por el valle del Nilo, llegó al Mediterráneo y, después, hacia el oeste y el norte, a Europa, y hacia el este a Oriente Medio, la India y el sudeste de Asia.
Plasmodium vivax es la especie que tiene una distribución más amplia, en el trópico, subtrópico y zonas templadas. P. falciparum es la especie más común en el trópico y subtrópicos y, rara vez, en zonas templadas. P. malariae tiene igual distribución que falciparum pero es menos común. Y P. ovale se encuentra en el África tropical y, esporádicamente, en otras zonas del planeta.
Referencias:
Bianucci, R. et al. 2007. Immunological evidence of Plasmodium falciparum infectyion in an Egyptian child mummy from the Early Dynastic Period. Journal of Archaeological Science doi: 10.106/j.jas.2007.11.019
Bray, R.S. & P.C.C. Garnham. 1982. The life-cycle of primate malaria parasites. British Medical Bulletin 38: 117-122.
Brockington, F. 1964. La salud en el mundo. EUDEBA. Buenos Aires. 403 pp.
Bruce-Chwatt, L.J. 1965. Paleogénesis and paleo-epidemiology of primate malaria. Bulletin of the WHO 32: 363-387.
Coluzzi, M. 1994. Malaria and the Afrotropical ecosystems: Impact of man-made environmental changes. Parasitologia 36: 223-227.
Earle, D.P. 1979. A history of malaria and its ironies. Transactions of the American Clinical and Climatological Association 90: 1-26.
Gentilini, M. et al. 1977. Médecine Tropicale. Flammarion-Sciences. Paris. 561 pp.
Hawas, Z. et al. 2010. Ancestry and pathology in King Tutankhemum’s family. Journal of American Medical Association 303: 638-647.
Hommel, M. 2007. Morphologie, biologie et cycle des Plasmodium parasites de l’homme. Bulletin de l’Académie nationale de médecine 191: 1235-1246.
Howes, R.E. et al. 2011. The global distribution of the Duffy blood group. Nature Communications DOI: 10.1038/ncomms1265
Laremruata, A. et al. 2013. Molecular identification of falciparum malaria and human tuberculosis co-infections in mummies from the Fayum Depression (Lower Egypt). PLOS ONE 8: e60307
Lepers, J.P.et al. 1990. Newly transmitted Plasmodium falciparum malaria in the central highland plateaux of Madagascar: assessment of clinical impact in a rural community. Bulletin of the WHO 68: 217-222.
Lu, W. Et al. 2014. African origins of the malaria parasite Plasmodium vivax. Nature Communications DOI: 10.1038/ncomms4346
Marciniak, S. et al. 2016. Plasmodium falciparum malaria in 1st-2nd century CE southern Italy. Current Biology DOI: 10.1016/j.cub.2016.10.016
Meibalan, E. & M. Marti. 2017. Biology of malaria transmission. Cold Spring Harbor Perspectives in Medicine 7: a025452
Mercereau, O. & D. Ménard. 2010. Plasmodium vivax and the Duffy antigen: A paradigm revisited. Transfusion Clinique et Biologique doi: 10.1016/j-tracli.2010.06.005
Pagès, F. 1953. Le paludisme. PUF. Paris. 113 pp.
Schlagenhauf, P. 2004. Malaria: from prehistory to present. Infectious Disease Clinics of North America 18: 189-205.
Tan, S.Y. & A. Ahana. 2009. Charles Laveran (1845-1922): Nobel laureate pioneer of malaria. Singapore Medical Journal 50: 657-658.
Sobre el autor: Eduardo Angulo es doctor en biología, profesor de biología celular de la UPV/EHU retirado y divulgador científico. Ha publicado varios libros y es autor de La biología estupenda.
El artículo Historias de la malaria: El parásito se ha escrito en Cuaderno de Cultura Científica.
Entradas relacionadas:- Historias de la malaria: El charlatán y caballero Sir Robert Talbor
- Historias de la malaria: El árbol de la quina
- Una buena colección de historias maravillosas
Migrantes nocturnos y radares meteorológicos
El primer estudio realizado en el Golfo de Bizkaia a gran escala sobre la migración nocturna de aves ha mostrado que la migración se realiza por el mar y tierra en primavera y por tierra en otoño, al contrario de lo que se creía. Para este trabajo, llevado a cabo entre la Sociedad de Ciencias Aranzadi y el Departamento de Matemáticas Aplicadas de la UPV/EHU, ha sido clave la utilización de radares meteorológicos como método de observación ornitológica.

Foto: Nadja Weisshaupt
La investigación realizada por Nadja Weisshaupt durante su tesis doctoral, pionera en el estudio de la migración nocturna en el Golfo de Bizkaia, rompe con varias creencias: “Se pensaba que la migración se realiza como un frente amplio en otoño, pero en mi estudio he visto que no es así”. Y es que, en cuanto a las épocas de migración, los resultados afirman que se registra una alta actividad nocturna en primavera y poca actividad en todo el otoño en Punta Galea. La migración otoñal se realiza más hacia los Pirineos, posiblemente evitando el cruce del mar. Por otro lado, las observaciones lunares han permitido establecer que las aves paseriformes (el grupo que abarca los pájaros cantores) predominan como migrantes nocturnos, aunque también se registra un pequeño porcentaje de aves acuáticas.
La migración de las aves incluye el desplazamiento de millones de individuos entre sus áreas de cría y los entornos a los que se desplazan para pasar el invierno. Durante sus migraciones, las aves recorren grandes distancias, para lo cual han de parar a menudo y reponer fuerzas para proseguir su viaje. Para poder alimentarse y descansar, necesitan ecosistemas adecuados en sus lugares de paso; la ausencia de entornos apropiados puede tener consecuencias tanto en la población como en la conservación de estas especies. En este escenario, el conocimiento de las estrategias migratorias es capital, tanto desde un punto de vista biológico y ecológico, como de la conservación. “Para proteger un espacio, es indispensable disponer de una base científica”, explica Nadja Weisshaupt, autora del estudio.
La migración de aves ha dado lugar a múltiples estudios científicos, en Euskadi sobre todo por anillamiento de aves capturadas, con el objeto de analizar los diversos factores que influyen en los patrones y dinámicas relacionados con la migración de las aves. “Sin embargo, hasta el momento se carecía de datos sobre la migración nocturna activa en el Golfo de Bizkaia”, añade. Con el objetivo de cubrir esta necesidad, partiendo de los conocimientos previos por anillamiento desde Aranzadi y la base de datos del radar de Punta Galea de Euskalmet, Weisshaupt ha abordado el estudio de la migración nocturna de aves en la costa del Golfo de Bizkaia.
Dadas las dificultades de visibilidad de noche y la necesidad de cubrir largas distancias, se recurrió a diversos métodos de observación: radares, cámara térmica y observaciones lunares.
Los radares meteorológicos constituyen la herramienta clave de este estudio. Se ha trabajado con dos tipos de radares. Uno de ellos es el perfilador de Euskalmet que se encuentra en Punta Galea. El perfilador realiza mediciones de viento, y en épocas de migración, registra señales que no corresponden a vientos, sino a las aves que pasan. Este hecho, que supone un gran problema para la calidad de los datos meteorológicos, resulta útil para el estudio de aves. “Es interesante, porque hemos tenido que luchar bastante para que la gente crea que es posible utilizar perfiladores para estudios ornitológicos”, señala Weisshaupt. El segundo tipo de radar mide la precipitación. El radar de Kapildui, por ejemplo, también de Euskalmet. La onda de este tipo de radar es más corta que la del perfilador, aunque también registra aves.
Pero no todos los radares son válidos; durante el estudio se ha constatado que los diferentes servicios meteorológicos se someten a diferentes niveles de procesamiento y filtrado de forma que los datos disponibles ya no contienen información biológica. Como consecuencia, se han descartado los radares de AEMET que hubiera cubierto la costa sur del golfo, pues solo los datos del radar de Kapildui de Euskalmet y de los radares franceses contienen información válida. Con estos cinco radares restantes se ha utilizado un algoritmo que permite automatizar la extracción de datos de aves de los radares meteorológicos, desarrollado por el doctor Adriaan Dokter de la Universidad de Ámsterdam, mediante una colaboración realizada dentro de la Acción COST ENRAM (European Network for the Radar Surveillance of Animal Movement) a nivel europeo.
El segundo tipo de herramienta, la cámara térmica, registra la radiación de infrarrojos que emiten varios objetos, por lo que detecta “la temperatura de un ave, 40-41 ºC, que contrasta claramente con la temperatura ambiente. La cámara permite especificar si las aves observadas son paseriformes, si van solas o en grupos. Además se obtiene la dirección y la intensidad de la migración del espacio de tiempo analizado”.
Finalmente, la observación lunar se realiza desde dos días antes hasta dos días después de la luna llena. “En este caso, se ha utilizado para complementar la información obtenida de los otros sistemas, a fin de especificar la composición de las aves migratorias”.
Referencia:
Nadja Weisshaupt, Mercedes Maruri, and Juan Arizaga (2014) Bird migration across the Bay of Biscay observed by meteorological radar [PDF] ERAD-14
Edición realizada por César Tomé López a partir de materiales suministrados por UPV/EHU Komunikazioa
El artículo Migrantes nocturnos y radares meteorológicos se ha escrito en Cuaderno de Cultura Científica.
Entradas relacionadas:- Algoritmos para que los parques eólicos no interfieran con los radares
- El aporte de amonio aumenta la producción de sustancias anticancerígenas en el brócoli
- Equilibrio tautomérico en un sistema modelo de gran interés biológico